

Molecular and Supramolecular Structure of the Mitochondrial Oxidative Phosphorylation System: Implications for Pathology
- PMID: 33804034
- PMCID: PMC7999509
- DOI: 10.3390/life11030242
Molecular and Supramolecular Structure of the Mitochondrial Oxidative Phosphorylation System: Implications for Pathology
Abstract
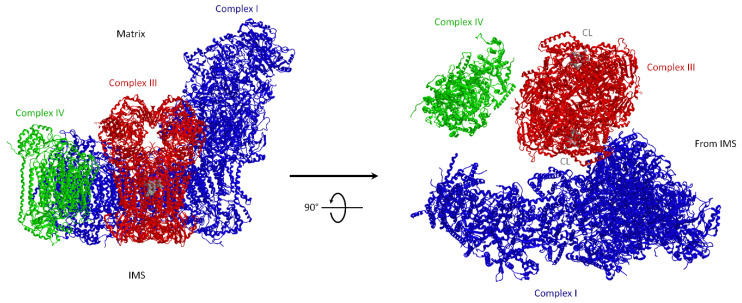
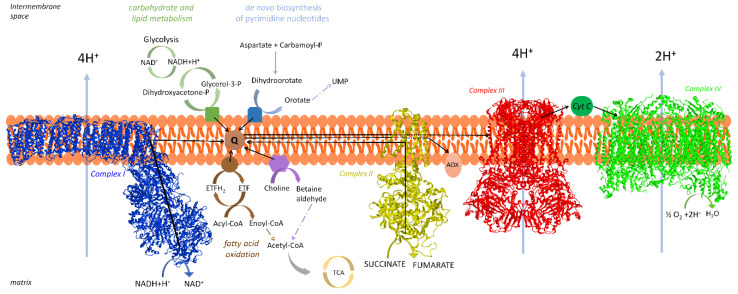
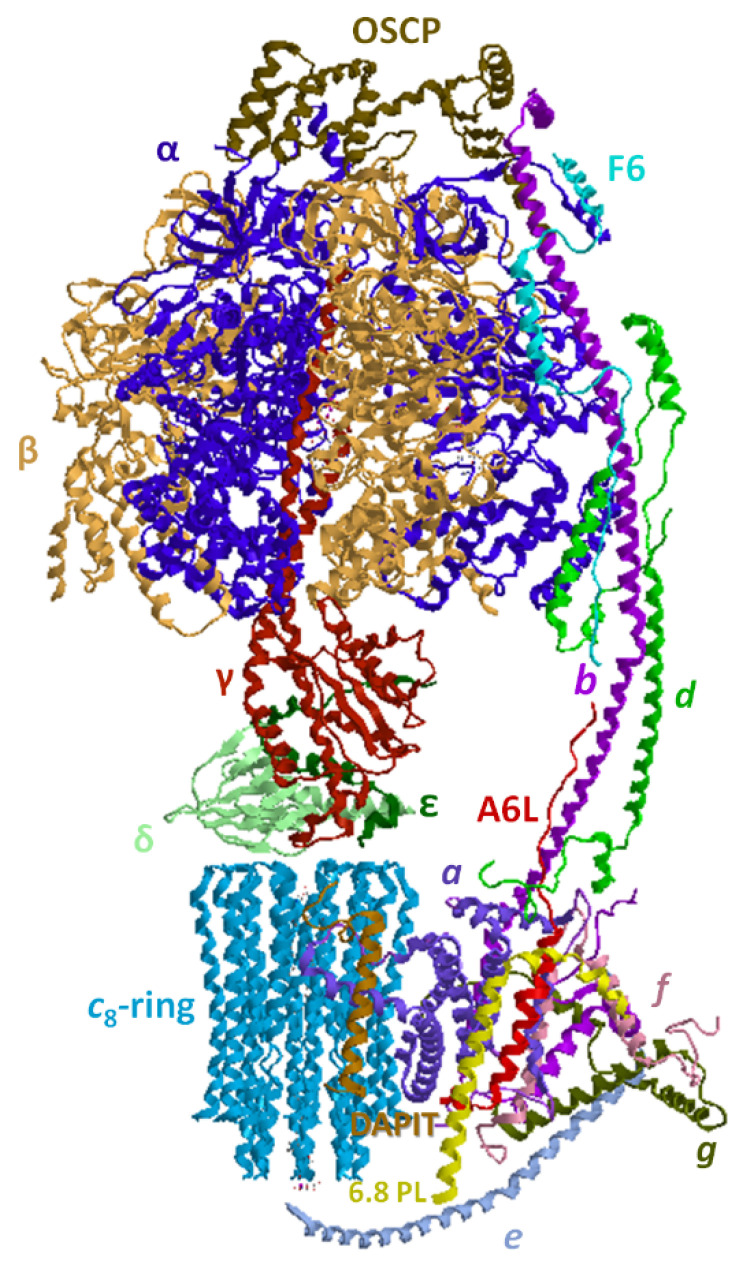
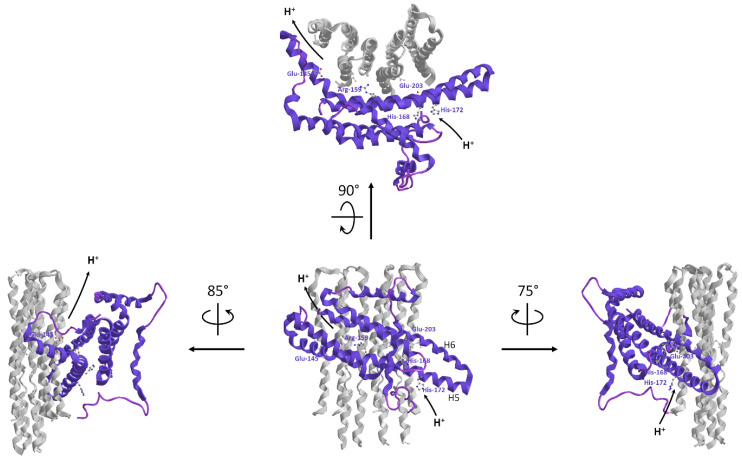
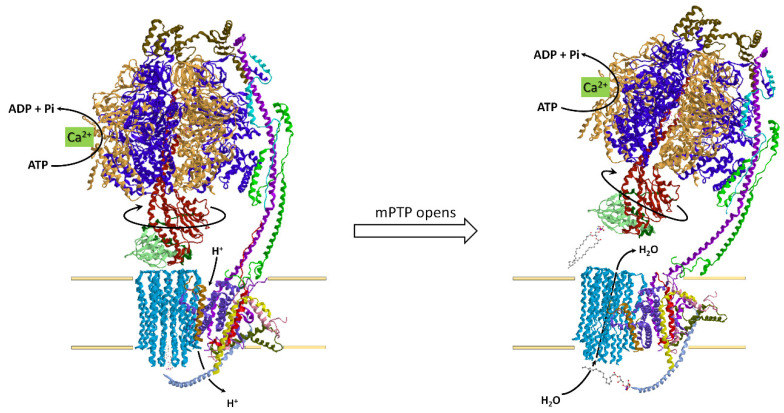
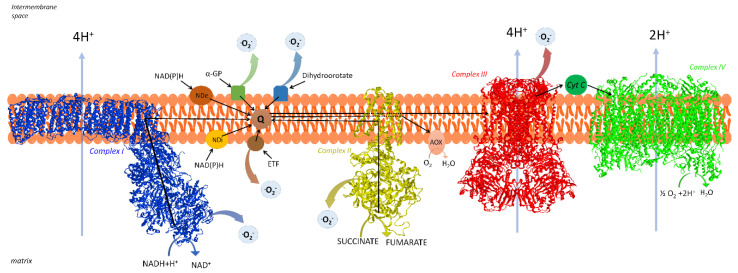
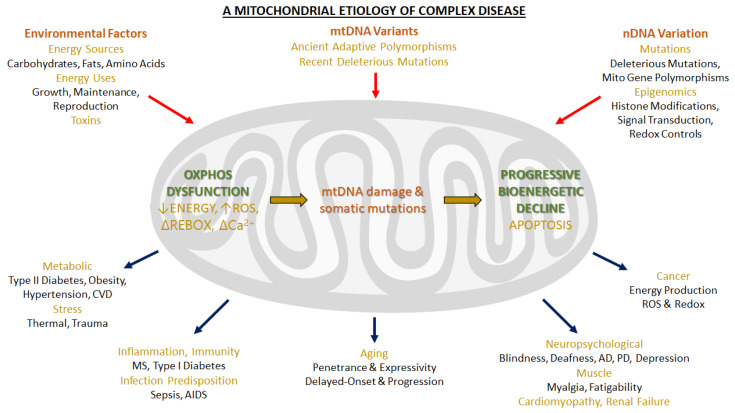
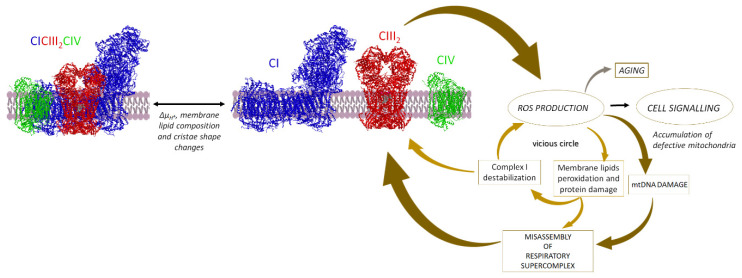
Under aerobic conditions, mitochondrial oxidative phosphorylation (OXPHOS) converts the energy released by nutrient oxidation into ATP, the currency of living organisms. The whole biochemical machinery is hosted by the inner mitochondrial membrane (mtIM) where the protonmotive force built by respiratory complexes, dynamically assembled as super-complexes, allows the F1FO-ATP synthase to make ATP from ADP + Pi. Recently mitochondria emerged not only as cell powerhouses, but also as signaling hubs by way of reactive oxygen species (ROS) production. However, when ROS removal systems and/or OXPHOS constituents are defective, the physiological ROS generation can cause ROS imbalance and oxidative stress, which in turn damages cell components. Moreover, the morphology of mitochondria rules cell fate and the formation of the mitochondrial permeability transition pore in the mtIM, which, most likely with the F1FO-ATP synthase contribution, permeabilizes mitochondria and leads to cell death. As the multiple mitochondrial functions are mutually interconnected, changes in protein composition by mutations or in supercomplex assembly and/or in membrane structures often generate a dysfunctional cascade and lead to life-incompatible diseases or severe syndromes. The known structural/functional changes in mitochondrial proteins and structures, which impact mitochondrial bioenergetics because of an impaired or defective energy transduction system, here reviewed, constitute the main biochemical damage in a variety of genetic and age-related diseases.
Keywords: ATP synthase/hydrolase; ROS; cellular signaling; cristae; mitochondrial dysfunction; mitochondrial permeability transition pore; oxidative phosphorylation; respiratory supercomplexes.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
