

Two Faces of Autophagy in the Struggle against Cancer
- PMID: 33804163
- PMCID: PMC8000091
- DOI: 10.3390/ijms22062981
Two Faces of Autophagy in the Struggle against Cancer
Abstract
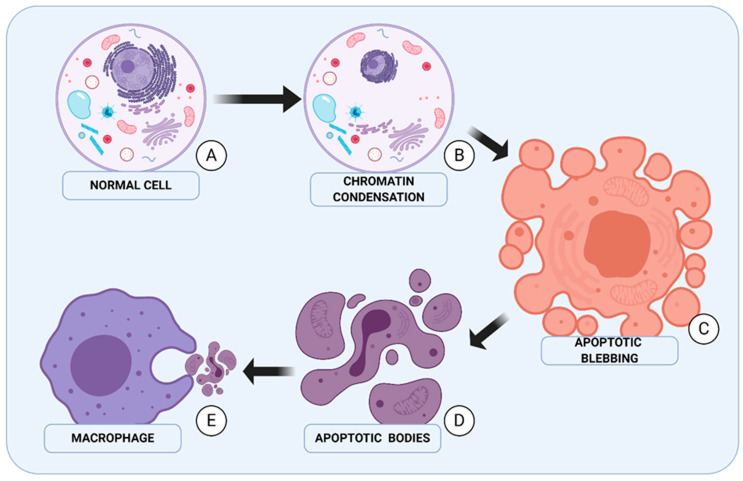
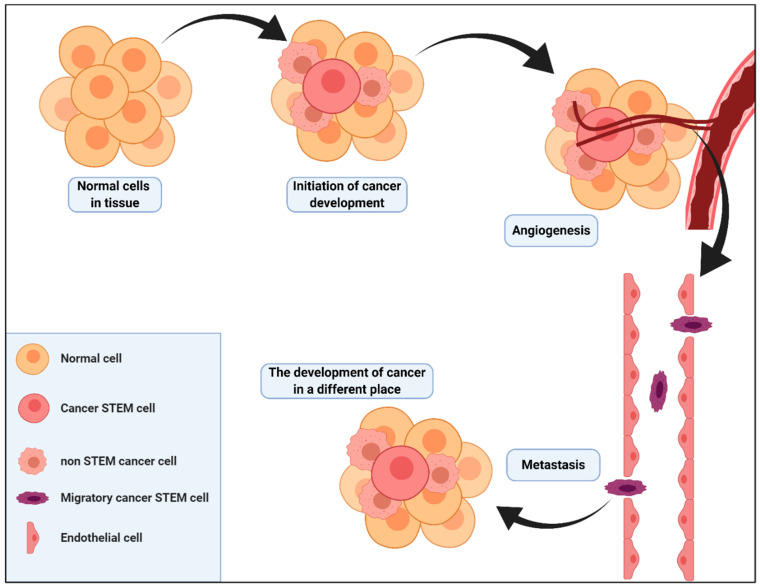
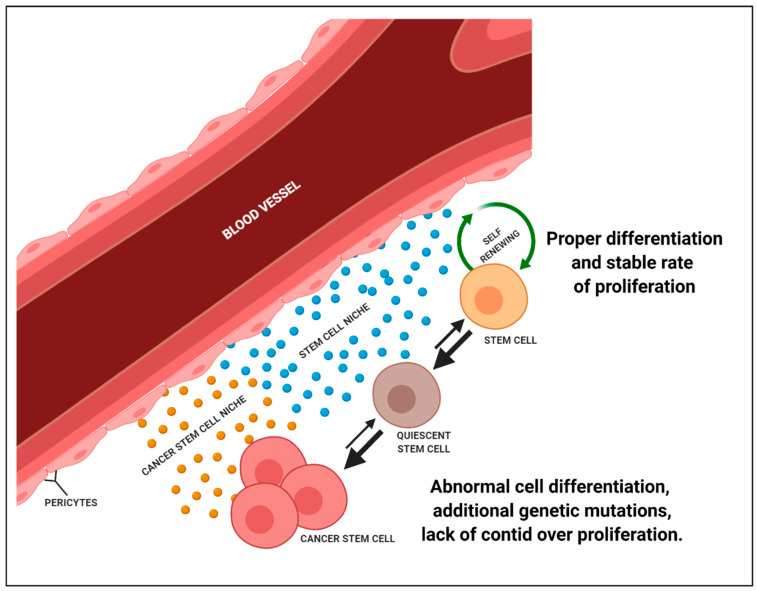
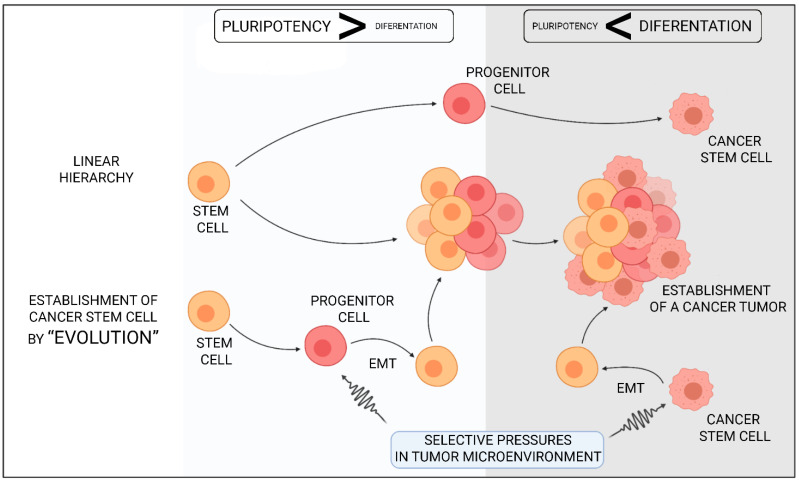
Autophagy can play a double role in cancerogenesis: it can either inhibit further development of the disease or protect cells, causing stimulation of tumour growth. This phenomenon is called "autophagy paradox", and is characterised by the features that the autophagy process provides the necessary substrates for biosynthesis to meet the cell's energy needs, and that the over-programmed activity of this process can lead to cell death through apoptosis. The fight against cancer is a difficult process due to high levels of resistance to chemotherapy and radiotherapy. More and more research is indicating that autophagy may play a very important role in the development of resistance by protecting cancer cells, which is why autophagy in cancer therapy can act as a "double-edged sword". This paper attempts to analyse the influence of autophagy and cancer stem cells on tumour development, and to compare new therapeutic strategies based on the modulation of these processes.
Keywords: anticancer therapies; apoptosis; autophagy modulation; cancer stem cells; cell death.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

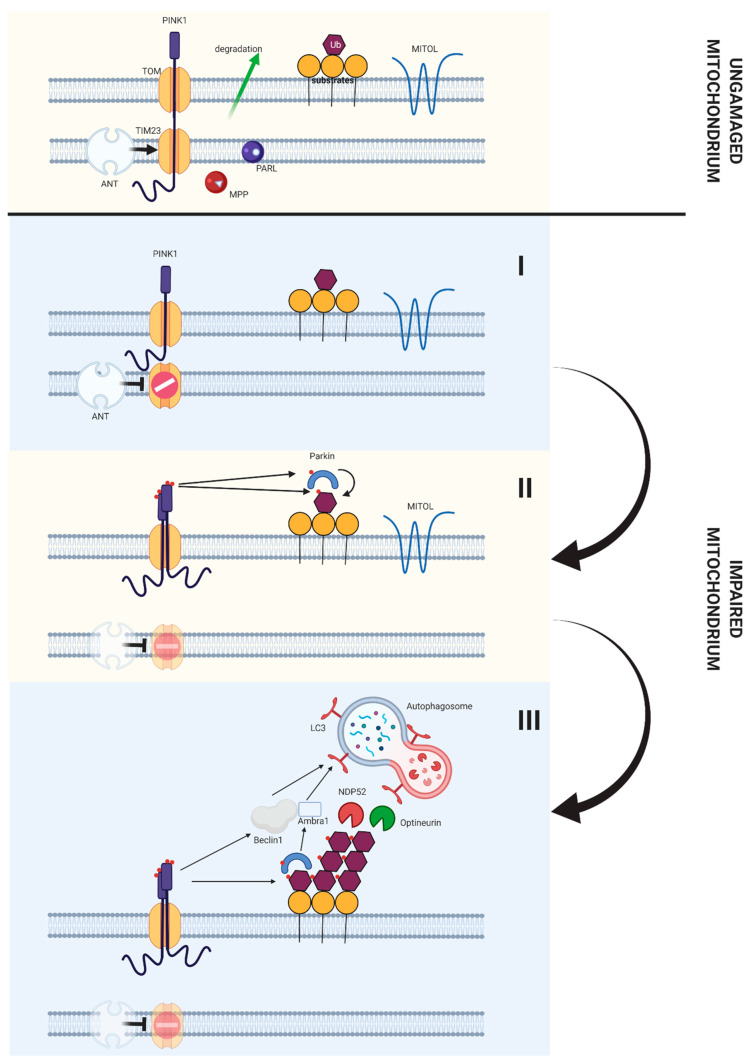
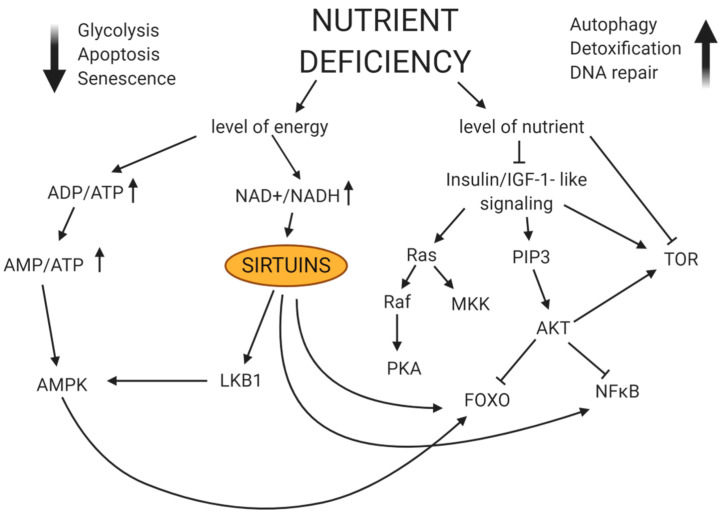
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
