

Islet Regeneration: Endogenous and Exogenous Approaches
- PMID: 33804882
- PMCID: PMC8037662
- DOI: 10.3390/ijms22073306
Islet Regeneration: Endogenous and Exogenous Approaches
Abstract
Both type 1 and type 2 diabetes are characterized by a progressive loss of beta cell mass that contributes to impaired glucose homeostasis. Although an optimal treatment option would be to simply replace the lost cells, it is now well established that unlike many other organs, the adult pancreas has limited regenerative potential. For this reason, significant research efforts are focusing on methods to induce beta cell proliferation (replication of existing beta cells), promote beta cell formation from alternative endogenous cell sources (neogenesis), and/or generate beta cells from pluripotent stem cells. In this article, we will review (i) endogenous mechanisms of beta cell regeneration during steady state, stress and disease; (ii) efforts to stimulate endogenous regeneration and transdifferentiation; and (iii) exogenous methods of beta cell generation and transplantation.
Keywords: beta cell; diabetes; islet; pancreas; regeneration.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures
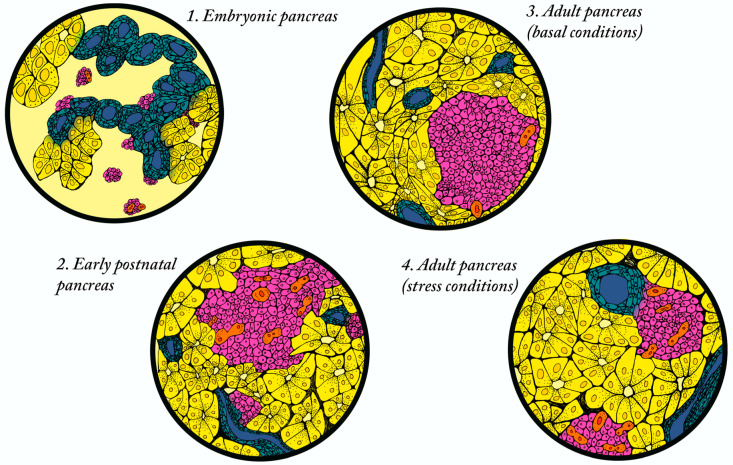
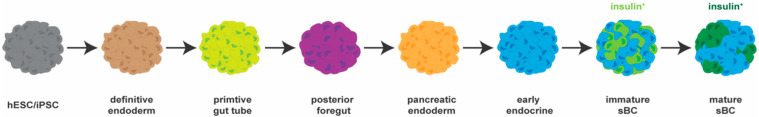
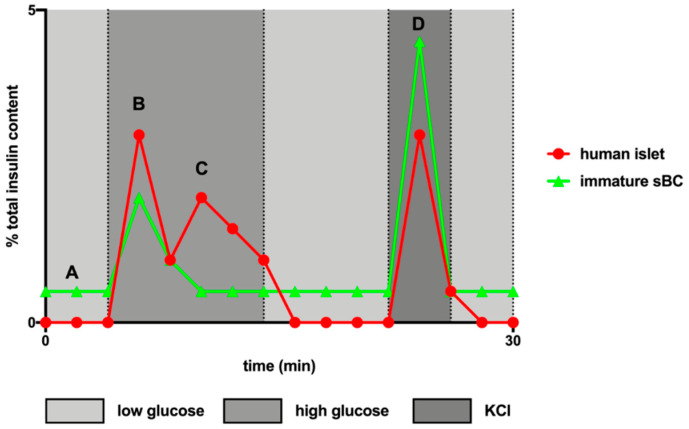
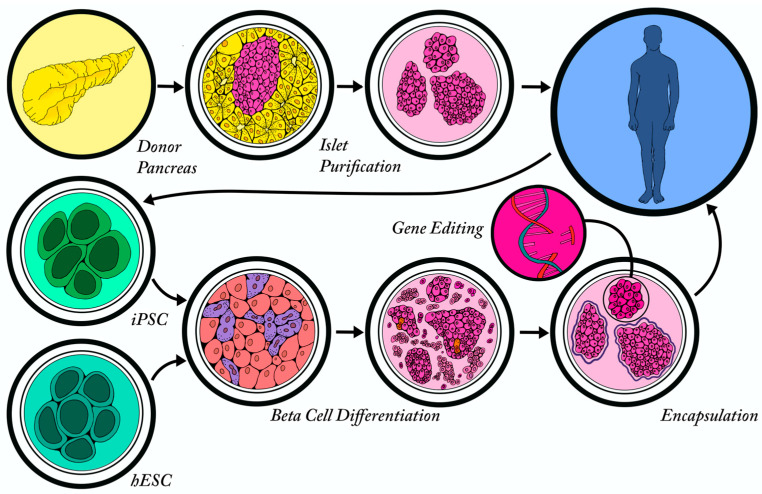
References
-
- Marselli L., Piron A., Suleiman M., Colli M.L., Yi X., Khamis A., Carrat G.R., Rutter G.A., Bugliani M., Giusti L., et al. Persistent or Transient Human beta Cell Dysfunction Induced by Metabolic Stress: Specific Signatures and Shared Gene Expression with Type 2 Diabetes. Cell Rep. 2020;33:108466. doi: 10.1016/j.celrep.2020.108466. - DOI - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
