

Adenovirus Terminal Protein Contains a Bipartite Nuclear Localisation Signal Essential for Its Import into the Nucleus
- PMID: 33804953
- PMCID: PMC8036708
- DOI: 10.3390/ijms22073310
Adenovirus Terminal Protein Contains a Bipartite Nuclear Localisation Signal Essential for Its Import into the Nucleus
Abstract
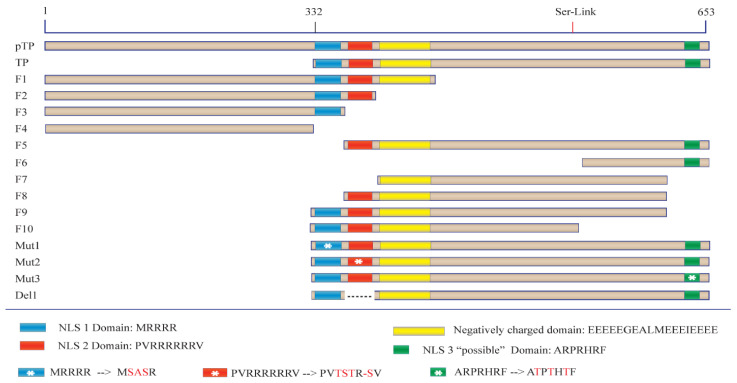
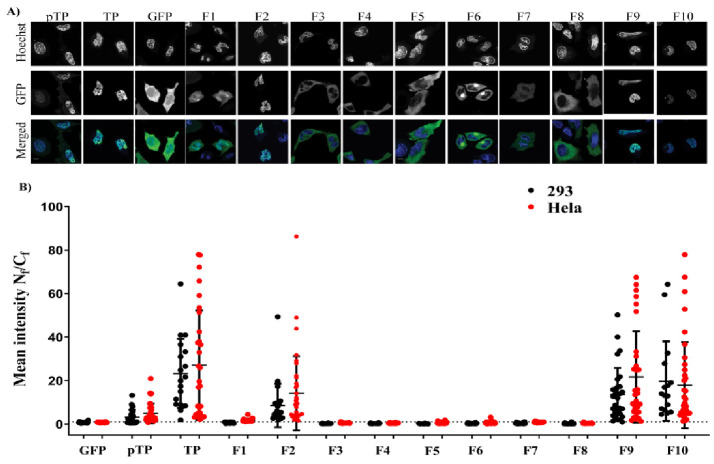
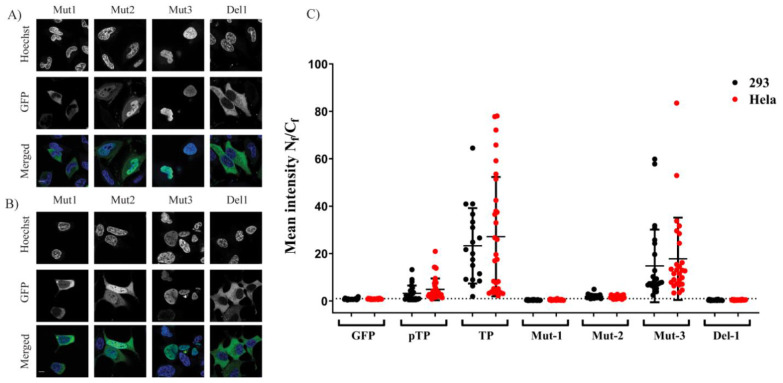
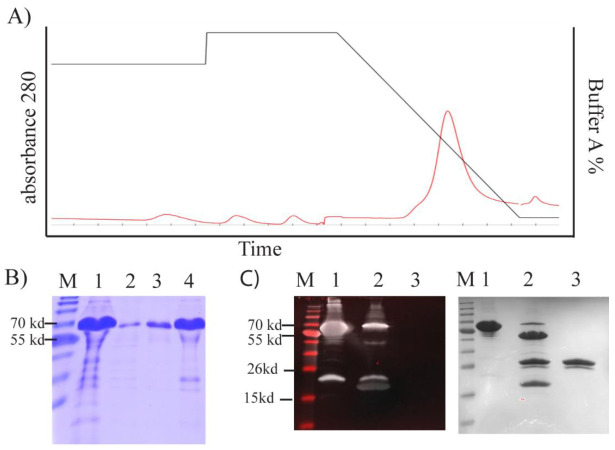
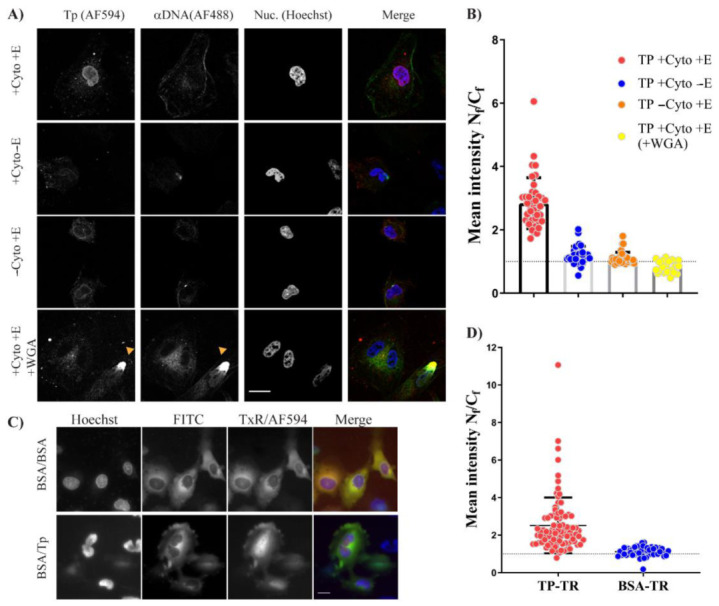
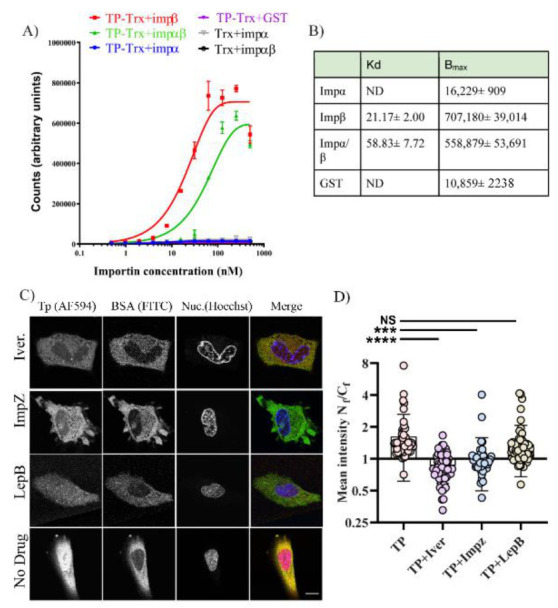
Adenoviruses contain dsDNA covalently linked to a terminal protein (TP) at the 5'end. TP plays a pivotal role in replication and long-lasting infectivity. TP has been reported to contain a nuclear localisation signal (NLS) that facilitates its import into the nucleus. We studied the potential NLS motifs within TP using molecular and cellular biology techniques to identify the motifs needed for optimum nuclear import. We used confocal imaging microscopy to monitor the localisation and nuclear association of GFP fusion proteins. We identified two nuclear localisation signals, PV(R)6VP and MRRRR, that are essential for fully efficient TP nuclear entry in transfected cells. To study TP-host interactions further, we expressed TP in Escherichia coli (E. coli). Nuclear uptake of purified protein was determined in digitonin-permeabilised cells. The data confirmed that nuclear uptake of TP requires active transport using energy and shuttling factors. This mechanism of nuclear transport was confirmed when expressed TP was microinjected into living cells. Finally, we uncovered the nature of TP binding to host nuclear shuttling proteins, revealing selective binding to Imp β, and a complex of Imp α/β but not Imp α alone. TP translocation to the nucleus could be inhibited using selective inhibitors of importins. Our results show that the bipartite NLS is required for fully efficient TP entry into the nucleus and suggest that this translocation can be carried out by binding to Imp β or Imp α/β. This work forms the biochemical foundation for future work determining the involvement of TP in nuclear delivery of adenovirus DNA.
Keywords: DNA binding proteins (DBP); DNA viruses; adenovirus; nuclear localisation signal (NLS); preterminal protein (pTP); terminal protein (TP); viral genome.
Conflict of interest statement
The authors declare no conflict of interest. The funders of H.A.A.-W.’s scholarship had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures

References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
