

The Telomeric Protein TRF2 Regulates Replication Origin Activity within Pericentromeric Heterochromatin
- PMID: 33804994
- PMCID: PMC8063955
- DOI: 10.3390/life11040267
The Telomeric Protein TRF2 Regulates Replication Origin Activity within Pericentromeric Heterochromatin
Abstract
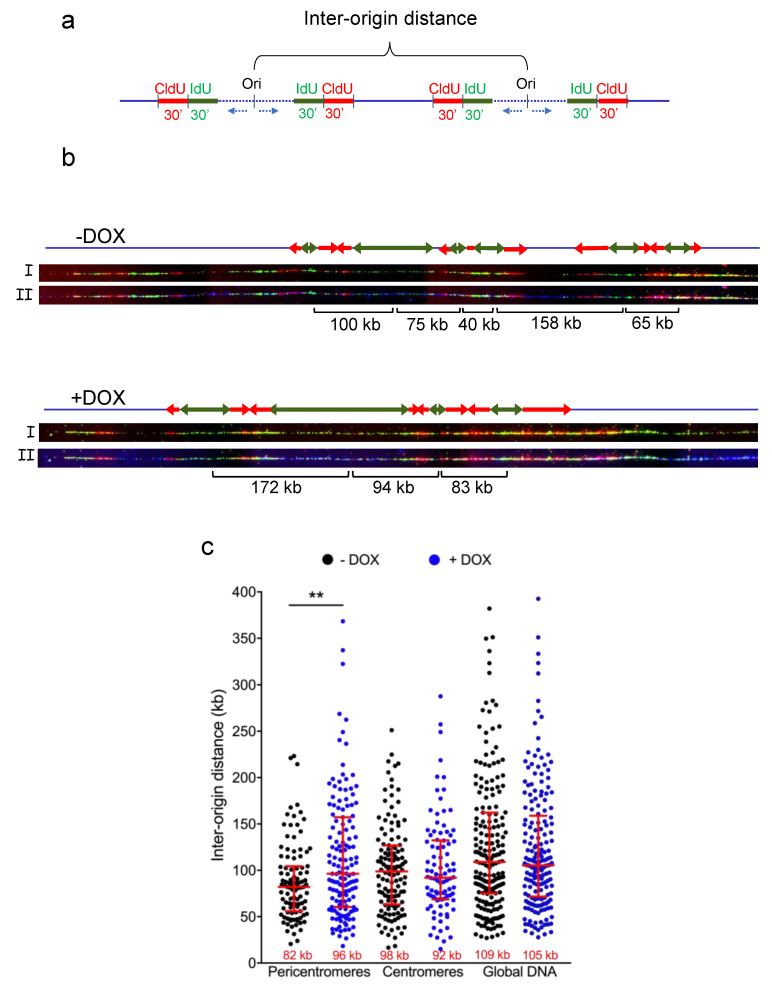
Heterochromatic regions render the replication process particularly difficult due to the high level of chromatin compaction and the presence of repeated DNA sequences. In humans, replication through pericentromeric heterochromatin requires the binding of a complex formed by the telomeric factor TRF2 and the helicase RTEL1 in order to relieve topological barriers blocking fork progression. Since TRF2 is known to bind the Origin Replication Complex (ORC), we hypothesized that this factor could also play a role at the replication origins (ORI) of these heterochromatin regions. By performing DNA combing analysis, we found that the ORI density is higher within pericentromeric satellite DNA repeats than within bulk genomic DNA and decreased upon TRF2 downregulation. Moreover, we showed that TRF2 and ORC2 interact in pericentromeric DNA, providing a mechanism by which TRF2 is involved in ORI activity. Altogether, our findings reveal an essential role for TRF2 in pericentromeric heterochromatin replication by regulating both replication initiation and elongation.
Keywords: ORC; TRF2; heterochromatin; pericentromeric DNA; replication; telomeres.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


References
-
- Tazumi A., Fukuura M., Nakato R., Kishimoto A., Takenaka T., Ogawa S., Song J.H., Takahashi T.S., Nakagawa T., Shirahige K., et al. Telomere-binding protein Taz1 controls global replication timing through its localization near late replication origins in fission yeast. Genes Dev. 2012;26:2050–2062. doi: 10.1101/gad.194282.112. - DOI - PMC - PubMed
Grants and funding
- ANR-11-LABX-0028-01/Institut National de la Santé et de la Recherche Médicale
- REPLITOP/Institut National Du Cancer
- TELOCHROM/Agence Nationale de la Recherche
- numbers 81971312, 91749126, 81911530241, 81871549 and 81671900/National Natural Science Foundation of China
- Oriental Scholars Program, 2019/Shanghai Municipal Education Commission
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
