

Conformational Heterogeneity and Cooperative Effects of Mammalian ALOX15
- PMID: 33807076
- PMCID: PMC8004969
- DOI: 10.3390/ijms22063285
Conformational Heterogeneity and Cooperative Effects of Mammalian ALOX15
Abstract
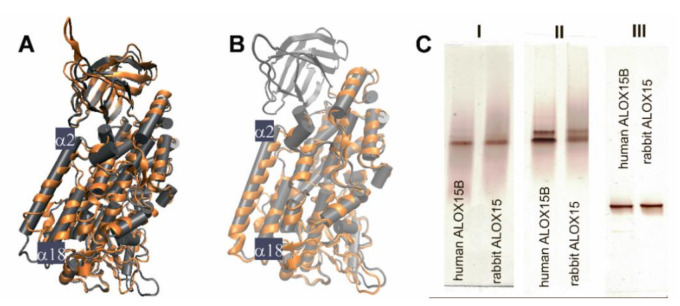
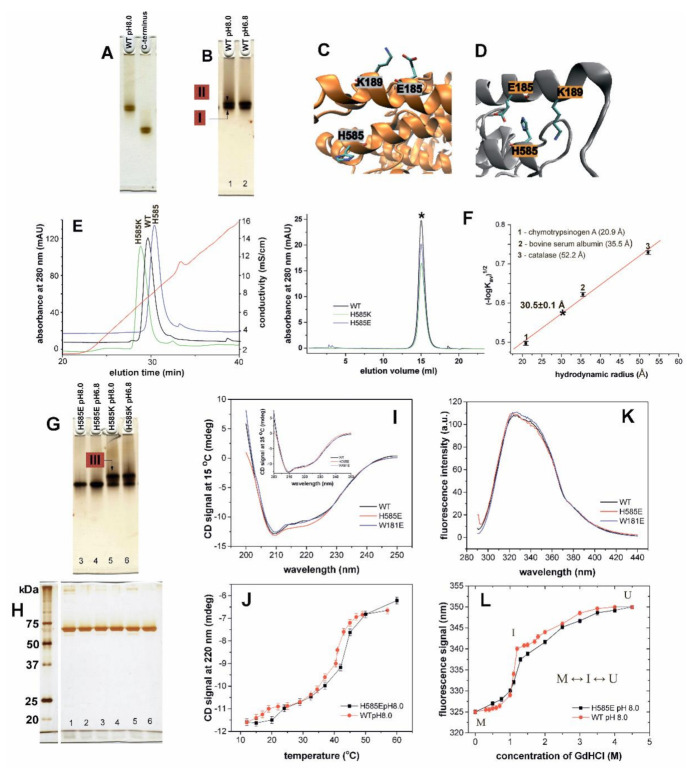
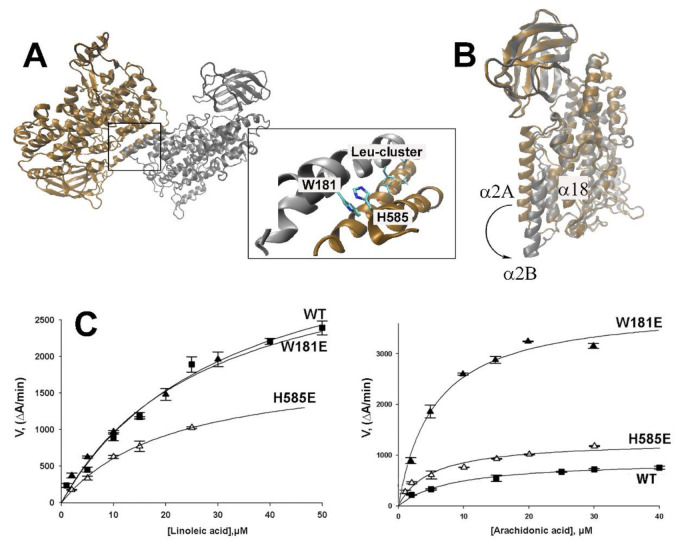
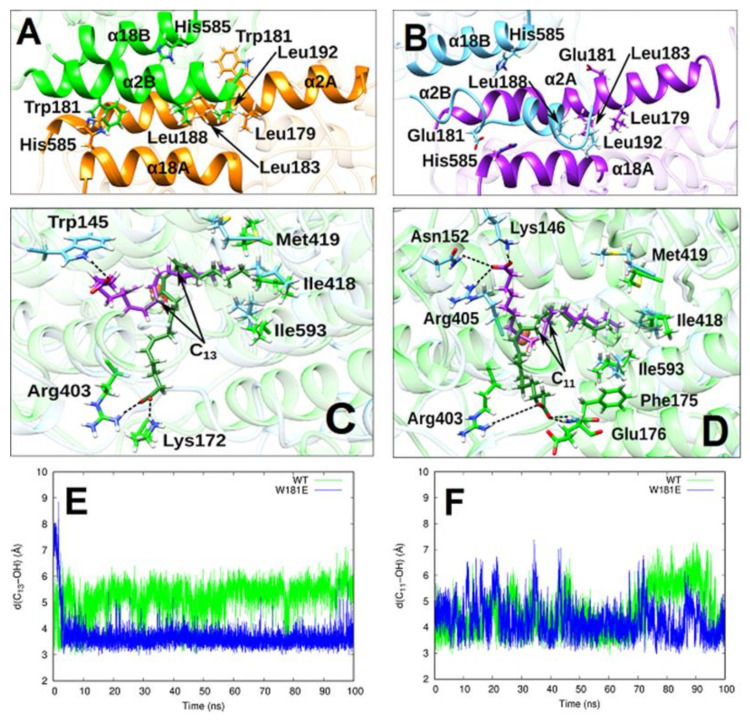
Arachidonic acid lipoxygenases (ALOXs) have been suggested to function as monomeric enzymes, but more recent data on rabbit ALOX15 indicated that there is a dynamic monomer-dimer equilibrium in aqueous solution. In the presence of an active site ligand (the ALOX15 inhibitor RS7) rabbit ALOX15 was crystalized as heterodimer and the X-ray coordinates of the two monomers within the dimer exhibit subtle structural differences. Using native polyacrylamide electrophoresis, we here observed that highly purified and predominantly monomeric rabbit ALOX15 and human ALOX15B are present in two conformers with distinct electrophoretic mobilities. In silico docking studies, molecular dynamics simulations, site directed mutagenesis experiments and kinetic measurements suggested that in aqueous solutions the two enzymes exhibit motional flexibility, which may impact the enzymatic properties.
Keywords: cooperative effects; crystal structure; lipoxygenases; molecular dynamics; protein–protein interactions.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study or in the decision to publish the results
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
