

Computational Analysis of the Interactions between the S100B Extracellular Chaperone and Its Amyloid β Peptide Client
- PMID: 33807304
- PMCID: PMC8037576
- DOI: 10.3390/ijms22073629
Computational Analysis of the Interactions between the S100B Extracellular Chaperone and Its Amyloid β Peptide Client
Abstract
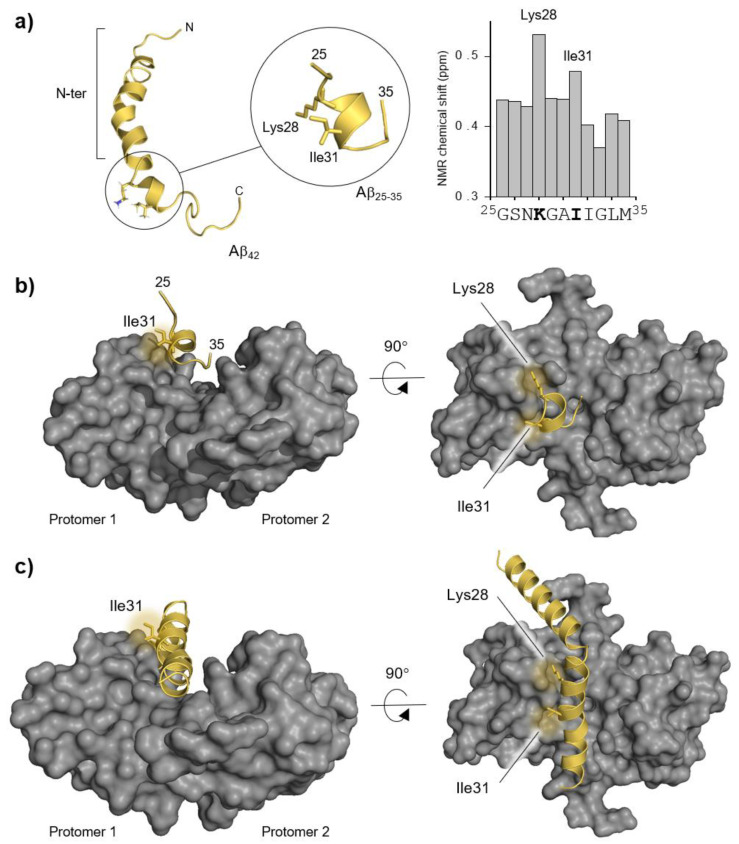
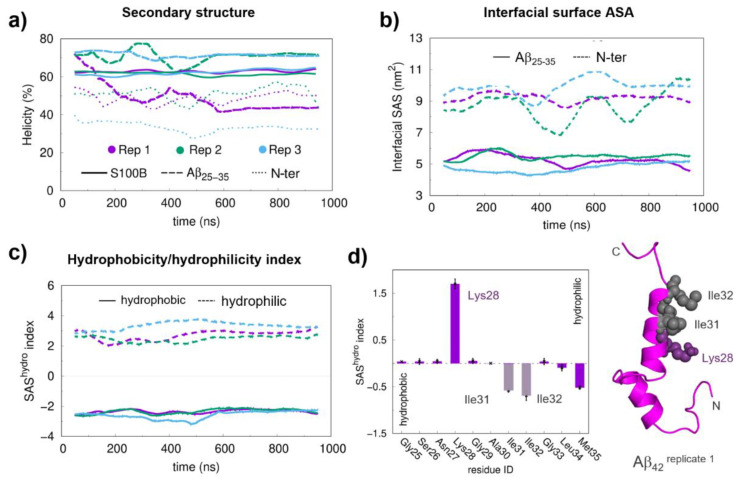
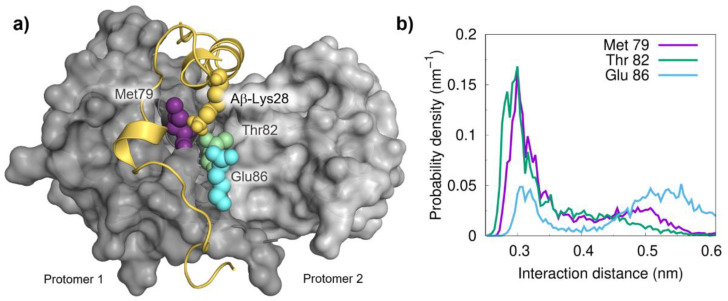
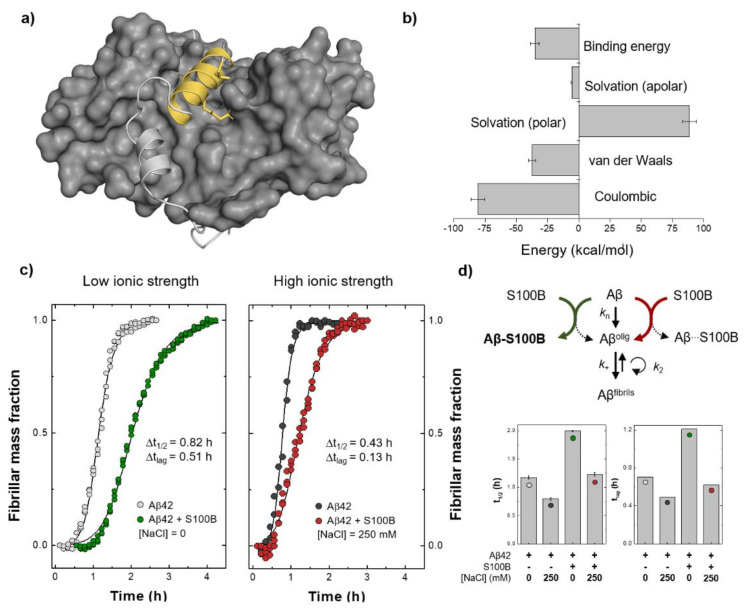
S100B is an astrocytic extracellular Ca2+-binding protein implicated in Alzheimer's disease, whose role as a holdase-type chaperone delaying Aβ42 aggregation and toxicity was recently uncovered. Here, we employ computational biology approaches to dissect the structural details and dynamics of the interaction between S100B and Aβ42. Driven by previous structural data, we used the Aβ25-35 segment, which recapitulates key aspects of S100B activity, as a starting guide for the analysis. We used Haddock to establish a preferred binding mode, which was studied with the full length Aβ using long (1 μs) molecular dynamics (MD) simulations to investigate the structural dynamics and obtain representative interaction complexes. From the analysis, Aβ-Lys28 emerged as a key candidate for stabilizing interactions with the S100B binding cleft, in particular involving a triad composed of Met79, Thr82 and Glu86. Binding constant calculations concluded that coulombic interactions, presumably implicating the Lys28(Aβ)/Glu86(S100B) pair, are very relevant for the holdase-type chaperone activity. To confirm this experimentally, we examined the inhibitory effect of S100B over Aβ aggregation at high ionic strength. In agreement with the computational predictions, we observed that electrostatic perturbation of the Aβ-S100B interaction decreases anti-aggregation activity. Altogether, these findings unveil features relevant in the definition of selectivity of the S100B chaperone, with implications in Alzheimer's disease.
Keywords: amyloids; chaperones; docking; molecular dynamics; protein aggregation; protein folding diseases; protein interactions.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Tetramerization of the S100B Chaperone Spawns a Ca2+ Independent Regulatory Surface that Enhances Anti-aggregation Activity and Client Specificity.J Mol Biol. 2022 Oct 15;434(19):167791. doi: 10.1016/j.jmb.2022.167791. Epub 2022 Aug 12. J Mol Biol. 2022. PMID: 35970403
-
The S100B Alarmin Is a Dual-Function Chaperone Suppressing Amyloid-β Oligomerization through Combined Zinc Chelation and Inhibition of Protein Aggregation.ACS Chem Neurosci. 2020 Sep 2;11(17):2753-2760. doi: 10.1021/acschemneuro.0c00392. Epub 2020 Aug 7. ACS Chem Neurosci. 2020. PMID: 32706972
-
Dynamic interactions and Ca2+-binding modulate the holdase-type chaperone activity of S100B preventing tau aggregation and seeding.Nat Commun. 2021 Nov 1;12(1):6292. doi: 10.1038/s41467-021-26584-2. Nat Commun. 2021. PMID: 34725360 Free PMC article.
-
Promotion and Inhibition of Amyloid-β Peptide Aggregation: Molecular Dynamics Studies.Int J Mol Sci. 2021 Feb 13;22(4):1859. doi: 10.3390/ijms22041859. Int J Mol Sci. 2021. PMID: 33668406 Free PMC article. Review.
-
Exploring the Alzheimer amyloid-β peptide conformational ensemble: A review of molecular dynamics approaches.Peptides. 2015 Jul;69:86-91. doi: 10.1016/j.peptides.2015.04.009. Epub 2015 Apr 20. Peptides. 2015. PMID: 25908410 Review.
Cited by
-
SMOC1 colocalizes with Alzheimer's disease neuropathology and delays Aβ aggregation.Res Sq [Preprint]. 2024 Nov 1:rs.3.rs-5229472. doi: 10.21203/rs.3.rs-5229472/v1. Res Sq. 2024. Update in: Acta Neuropathol. 2024 Nov 25;148(1):72. doi: 10.1007/s00401-024-02819-6. PMID: 39574902 Free PMC article. Updated. Preprint.
-
Secondary Modification of S100B Influences Anti Amyloid-β Aggregation Activity and Alzheimer's Disease Pathology.Int J Mol Sci. 2024 Feb 1;25(3):1787. doi: 10.3390/ijms25031787. Int J Mol Sci. 2024. PMID: 38339064 Free PMC article.
-
Amyloid-β Processing in Aged S100B Transgenic Mice Is Sex Dependent.Int J Mol Sci. 2021 Oct 6;22(19):10823. doi: 10.3390/ijms221910823. Int J Mol Sci. 2021. PMID: 34639161 Free PMC article.
-
S100B chaperone multimers suppress the formation of oligomers during Aβ42 aggregation.Front Neurosci. 2023 Mar 21;17:1162741. doi: 10.3389/fnins.2023.1162741. eCollection 2023. Front Neurosci. 2023. PMID: 37025373 Free PMC article.
-
SMOC1 colocalizes with Alzheimer's disease neuropathology and delays Aβ aggregation.Acta Neuropathol. 2024 Nov 25;148(1):72. doi: 10.1007/s00401-024-02819-6. Acta Neuropathol. 2024. PMID: 39585417 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
