

Intracellular Toxic AGEs (TAGE) Triggers Numerous Types of Cell Damage
- PMID: 33808036
- PMCID: PMC8001776
- DOI: 10.3390/biom11030387
Intracellular Toxic AGEs (TAGE) Triggers Numerous Types of Cell Damage
Abstract
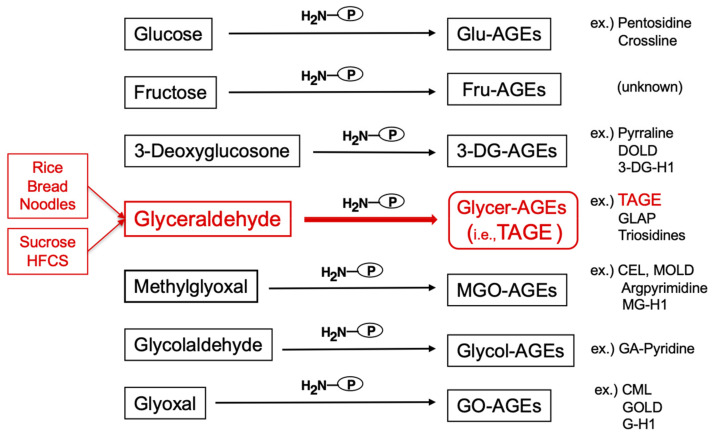
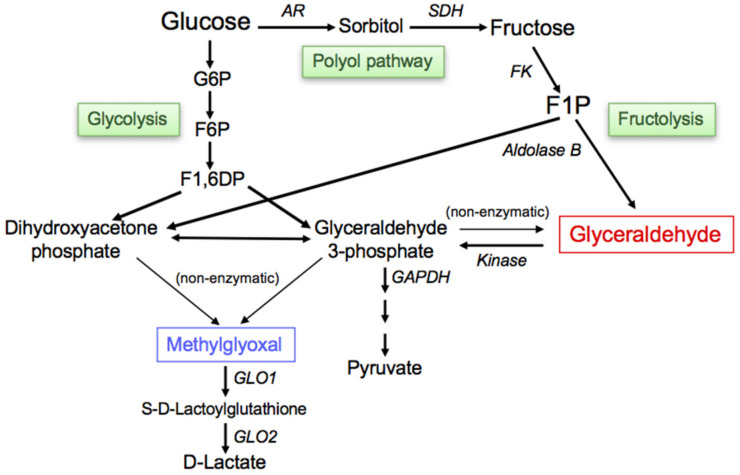
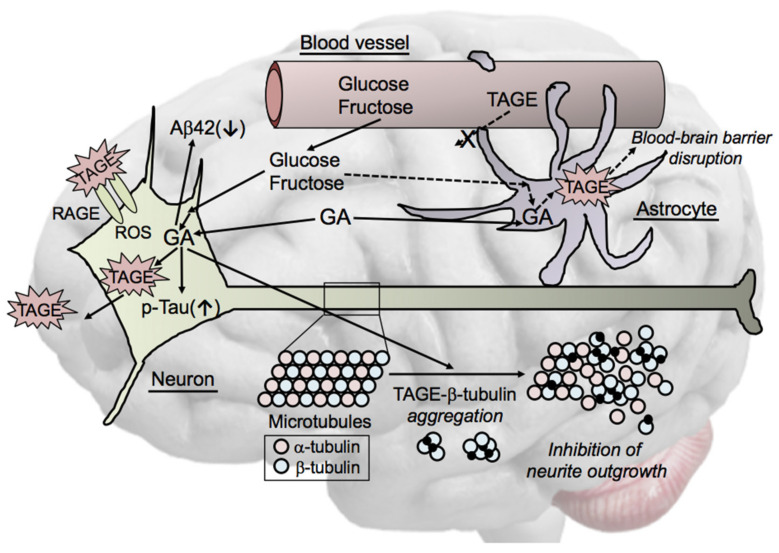
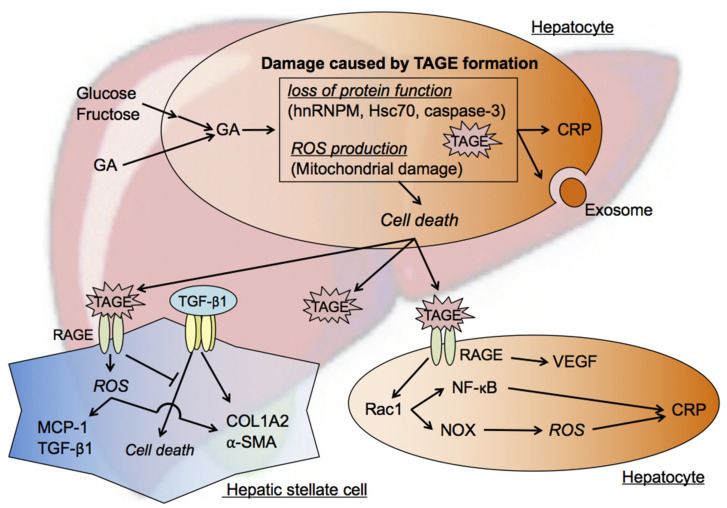
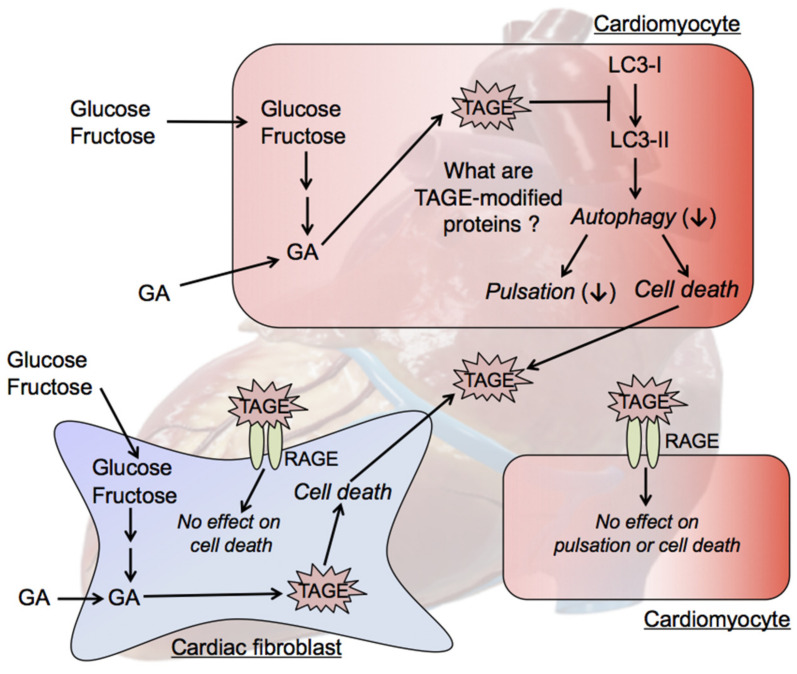
The habitual intake of large amounts of sugar, which has been implicated in the onset/progression of lifestyle-related diseases (LSRD), induces the excessive production of glyceraldehyde (GA), an intermediate of sugar metabolism, in neuronal cells, hepatocytes, and cardiomyocytes. Reactions between GA and intracellular proteins produce toxic advanced glycation end-products (toxic AGEs, TAGE), the accumulation of which contributes to various diseases, such as Alzheimer's disease, non-alcoholic steatohepatitis, and cardiovascular disease. The cellular leakage of TAGE affects the surrounding cells via the receptor for AGEs (RAGE), thereby promoting the onset/progression of LSRD. We demonstrated that the intracellular accumulation of TAGE triggered numerous cellular disorders, and also that TAGE leaked into the extracellular space, thereby increasing extracellular TAGE levels in circulating fluids. Intracellular signaling and the production of reactive oxygen species are affected by extracellular TAGE and RAGE interactions, which, in turn, facilitate the intracellular generation of TAGE, all of which may contribute to the pathological changes observed in LSRD. In this review, we discuss the relationships between intracellular TAGE levels and numerous types of cell damage. The novel concept of the "TAGE theory" is expected to open new perspectives for research into LSRD.
Keywords: Alzheimer’s disease (AD); advanced glycation end-products (AGEs); cardiovascular disease (CVD); lifestyle-related diseases (LSRD); non-alcoholic steatohepatitis (NASH); toxic AGEs (TAGE).
Conflict of interest statement
The authors declare that no conflict of interest exist.
Figures
References
-
- Saeedi P., Petersohn I., Salpea P., Malanda B., Karuranga S., Unwin N., Colagiuri S., Guariguata L., Motala A.A., Ogurtsova K., et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9th edition. Diabetes Res. Clin. Pract. 2019;157:107843. doi: 10.1016/j.diabres.2019.107843. - DOI - PubMed
-
- Imamura F., O’Connor L., Ye Z., Mursu J., Hayashino Y., Bhupathiraju S.N., Forouhi N.G. Consumption of sugar sweetened beverages, artificially sweetened beverages, and fruit juice and incidence of type 2 diabetes: Systematic review, meta-analysis, and estimation of population attributable fraction. Br. J. Sports Med. 2016;50:496–504. doi: 10.1136/bjsports-2016-h3576rep. - DOI - PMC - PubMed
-
- Schwingshackl L., Hoffmann G., Lampousi A.M., Knüppel S., Iqbal K., Schwedhelm C., Bechthold A., Schlesinger S., Boeing H. Food groups and risk of type 2 diabetes mellitus: A systematic review and meta-analysis of prospective studies. Eur. J. Epidemiol. 2017;32:363–375. doi: 10.1007/s10654-017-0246-y. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
