

Mechanisms of Proteolytic Enzymes and Their Inhibition in QM/MM Studies
- PMID: 33810118
- PMCID: PMC8004986
- DOI: 10.3390/ijms22063232
Mechanisms of Proteolytic Enzymes and Their Inhibition in QM/MM Studies
Abstract
Experimental evidence for enzymatic mechanisms is often scarce, and in many cases inadvertently biased by the employed methods. Thus, apparently contradictory model mechanisms can result in decade long discussions about the correct interpretation of data and the true theory behind it. However, often such opposing views turn out to be special cases of a more comprehensive and superior concept. Molecular dynamics (MD) and the more advanced molecular mechanical and quantum mechanical approach (QM/MM) provide a relatively consistent framework to treat enzymatic mechanisms, in particular, the activity of proteolytic enzymes. In line with this, computational chemistry based on experimental structures came up with studies on all major protease classes in recent years; examples of aspartic, metallo-, cysteine, serine, and threonine protease mechanisms are well founded on corresponding standards. In addition, experimental evidence from enzyme kinetics, structural research, and various other methods supports the described calculated mechanisms. One step beyond is the application of this information to the design of new and powerful inhibitors of disease-related enzymes, such as the HIV protease. In this overview, a few examples demonstrate the high potential of the QM/MM approach for sophisticated pharmaceutical compound design and supporting functions in the analysis of biomolecular structures.
Keywords: enzymes; proteases; qm/mm; quantum chemistry.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
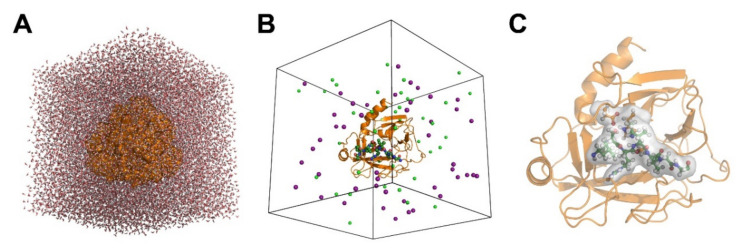
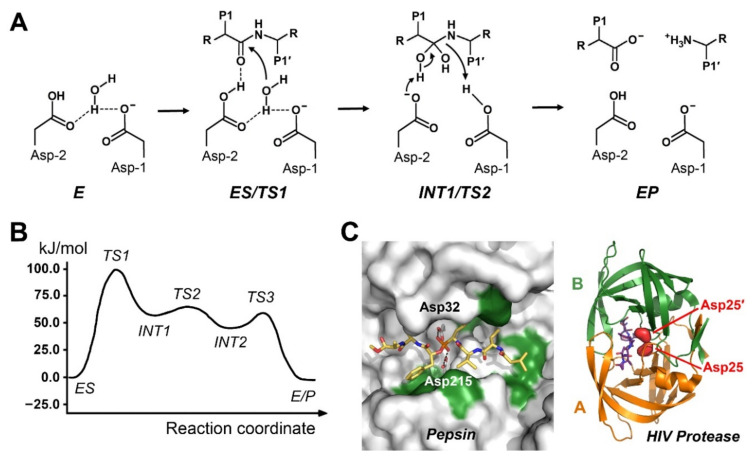
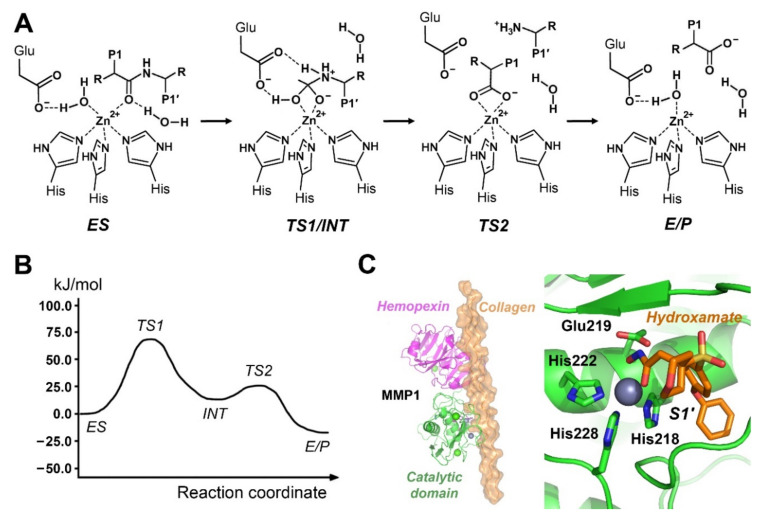
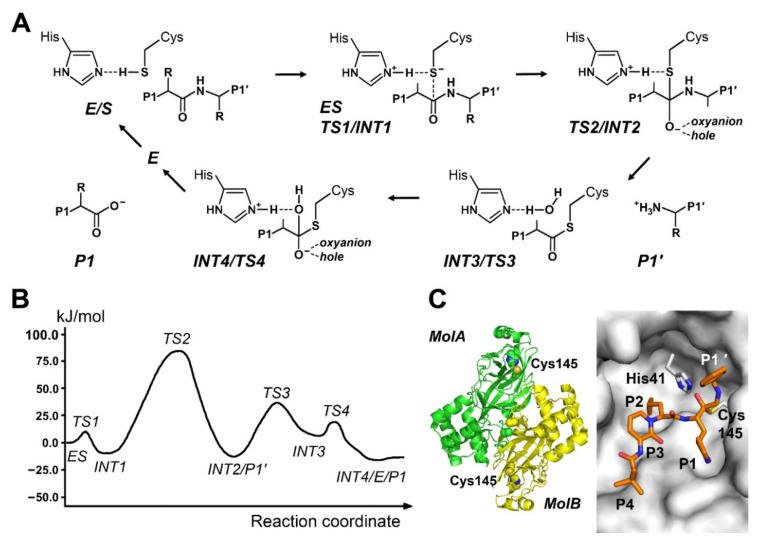
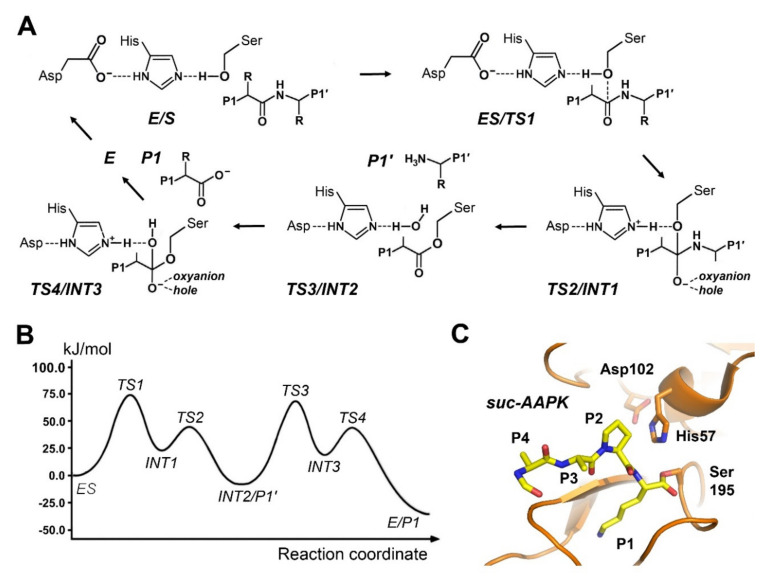
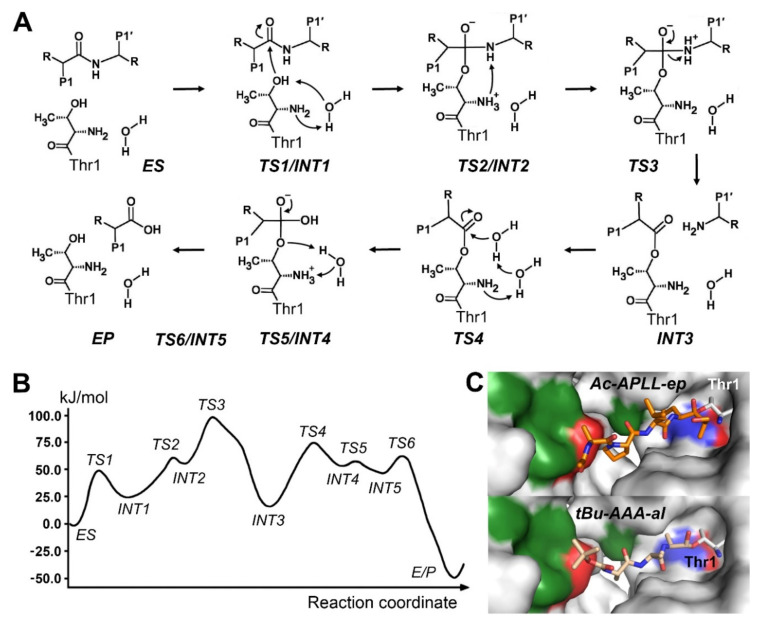
References
-
- Levene P.A. The Cleavage Products of Proteoses. J. Biol. Chem. 1905;1:45–58. doi: 10.1016/S0021-9258(17)46095-9. - DOI
-
- Robertson T.B. Note on the Synthesis of A Protein through the Action of Pepsin. J. Biol. Chem. 1907;3:95–99. doi: 10.1016/S0021-9258(17)45998-9. - DOI
-
- Bernal J.D., Crowfoot D. X-Ray Photographs of Crystalline Pepsin. Nature. 1934;133:794–795. doi: 10.1038/133794b0. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
