

Cross-Talk between the Complement Pathway and the Contact Activation System of Coagulation: Activated Factor XI Neutralizes Complement Factor H
- PMID: 33811105
- PMCID: PMC8030746
- DOI: 10.4049/jimmunol.2000398
Cross-Talk between the Complement Pathway and the Contact Activation System of Coagulation: Activated Factor XI Neutralizes Complement Factor H
Abstract
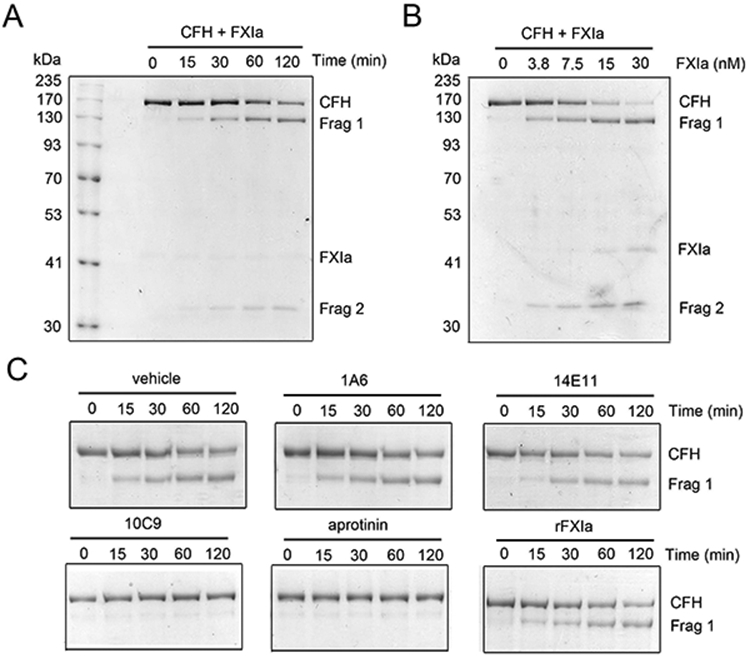
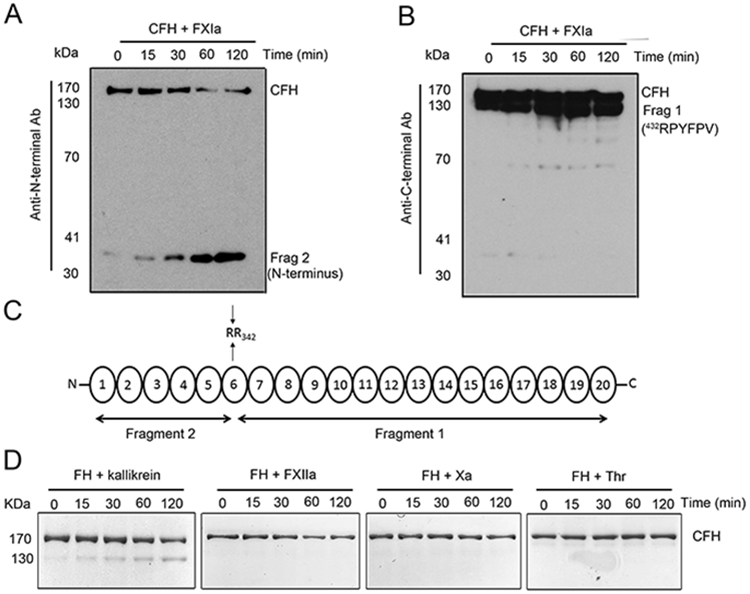
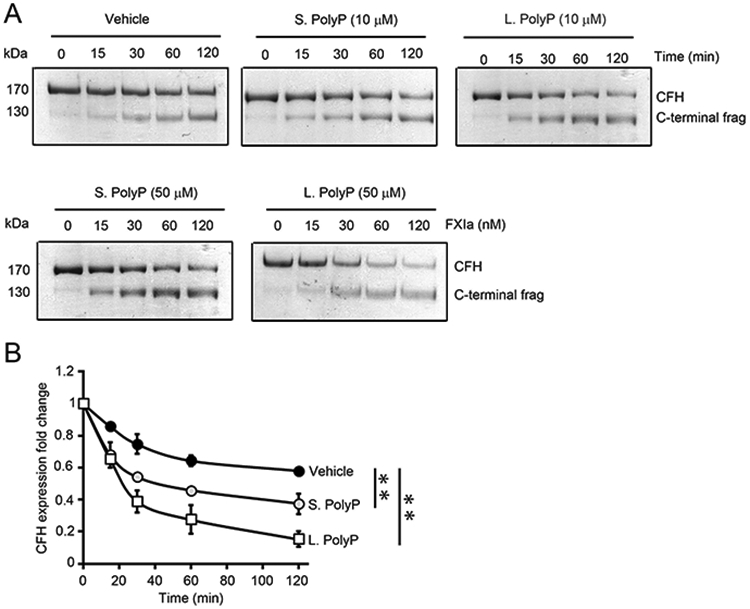
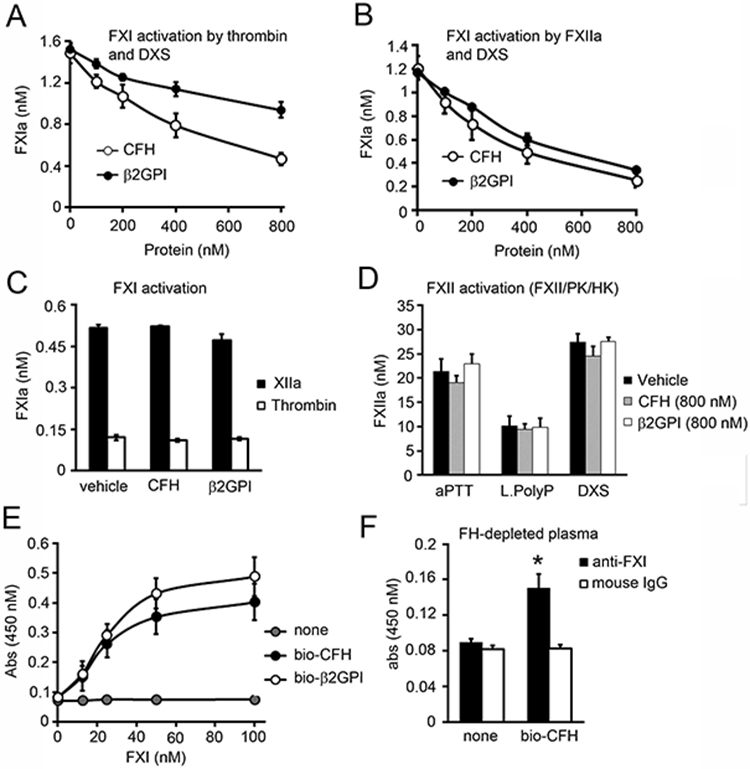
Complement factor H (CFH) is the major inhibitor of the alternative pathway of the complement system and is structurally related to beta2-glycoprotein I, which itself is known to bind to ligands, including coagulation factor XI (FXI). We observed reduced complement activation when FXI activation was inhibited in a baboon model of lethal systemic inflammation, suggesting cross-talk between FXI and the complement cascade. It is unknown whether FXI or its activated form, activated FXI (FXIa), directly interacts with the complement system. We explored whether FXI could interact with and inhibit the activity of CFH. We found that FXIa neutralized CFH by cleavage of the R341/R342 bonds. FXIa reduced the capacity of CFH to enhance the cleavage of C3b by factor I and the decay of C3bBb. The binding of CFH to human endothelial cells was also reduced after incubating CFH with FXIa. The addition of either short- or long-chain polyphosphate enhanced the capacity of FXIa to cleave CFH. FXIa also cleaved CFH that was present on endothelial cells and in the secretome from blood platelets. The generation of FXIa in plasma induced the cleavage of CFH. Moreover, FXIa reduced the cleavage of C3b by factor I in serum. Conversely, we observed that CFH inhibited FXI activation by either thrombin or FXIIa. Our study provides, to our knowledge, a novel molecular link between the contact pathway of coagulation and the complement system. These results suggest that FXIa generation enhances the activity of the complement system and thus may potentiate the immune response.
Copyright © 2021 by The American Association of Immunologists, Inc.
Conflict of interest statement
Disclosers
A. Gruber, C.U. Lorentz, E.I. Tucker, and Oregon Health & Science University have a significant financial interest in Aronora, Inc., a company that may have a commercial interest in the results of this research. This potential conflict of interest has been reviewed and managed by the Oregon Health & Science University Conflict of Interest in Research Committee. The remaining authors declare no competing financial interests.
Figures


Similar articles
-
Role of platelets in regulating activated coagulation factor XI activity.Am J Physiol Cell Physiol. 2021 Mar 1;320(3):C365-C374. doi: 10.1152/ajpcell.00056.2020. Epub 2021 Jan 20. Am J Physiol Cell Physiol. 2021. PMID: 33471623 Free PMC article.
-
Coagulation factor XI regulates endothelial cell permeability and barrier function in vitro and in vivo.Blood. 2024 Oct 24;144(17):1821-1833. doi: 10.1182/blood.2023022257. Blood. 2024. PMID: 39158072
-
Activated factor XI increases the procoagulant activity of the extrinsic pathway by inactivating tissue factor pathway inhibitor.Blood. 2015 Feb 26;125(9):1488-96. doi: 10.1182/blood-2014-10-604587. Epub 2015 Jan 13. Blood. 2015. PMID: 25587039 Free PMC article.
-
Substrates, Cofactors, and Cellular Targets of Coagulation Factor XIa.Semin Thromb Hemost. 2024 Oct;50(7):962-969. doi: 10.1055/s-0043-1764469. Epub 2023 Mar 20. Semin Thromb Hemost. 2024. PMID: 36940715 Free PMC article. Review.
-
The mechanism underlying activation of factor IX by factor XIa.Thromb Res. 2014 May;133 Suppl 1(0 1):S48-51. doi: 10.1016/j.thromres.2014.03.020. Thromb Res. 2014. PMID: 24759143 Free PMC article. Review.
Cited by
-
Activation of coagulation FXI promotes endothelial inflammation and amplifies platelet activation in a nonhuman primate model of hyperlipidemia.Res Pract Thromb Haemost. 2023 Nov 27;8(1):102276. doi: 10.1016/j.rpth.2023.102276. eCollection 2024 Jan. Res Pract Thromb Haemost. 2023. PMID: 38226339 Free PMC article.
-
Factor XI as a therapeutic target in neuroinflammatory disease.Curr Opin Hematol. 2024 Jan 1;31(1):32-38. doi: 10.1097/MOH.0000000000000787. Epub 2023 Sep 5. Curr Opin Hematol. 2024. PMID: 37694771 Free PMC article. Review.
-
Recombinant C1 inhibitor in the prevention of severe COVID-19: a randomized, open-label, multi-center phase IIa trial.Front Immunol. 2023 Oct 27;14:1255292. doi: 10.3389/fimmu.2023.1255292. eCollection 2023. Front Immunol. 2023. PMID: 37965347 Free PMC article. Clinical Trial.
-
Targeting thromboinflammation in COVID-19 - A narrative review of the potential of C1 inhibitor to prevent disease progression.Mol Immunol. 2022 Oct;150:99-113. doi: 10.1016/j.molimm.2022.08.008. Epub 2022 Aug 22. Mol Immunol. 2022. PMID: 36030710 Free PMC article. Review.
-
Oxidative Stress and Mitochondrial Damage in Dry Age-Related Macular Degeneration Like NFE2L2/PGC-1α -/- Mouse Model Evoke Complement Component C5a Independent of C3.Biology (Basel). 2021 Jul 4;10(7):622. doi: 10.3390/biology10070622. Biology (Basel). 2021. PMID: 34356477 Free PMC article.
References
-
- Sjöberg AP, Trouw LA, and Blom AM. 2009. Complement activation and inhibition: a delicate balance. Trends Immunol. 30: 83–90. - PubMed
-
- Perkins SJ, Nan R, Li K, Khan S, and Miller A. 2012. Complement Factor H-ligand interactions: Self-association, multivalency and dissociation constants. Immunobiology 217: 281–297. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
