

Paradoxical relationships between active transport and global protein distributions in neurons
- PMID: 33812847
- PMCID: PMC8390833
- DOI: 10.1016/j.bpj.2021.02.048
Paradoxical relationships between active transport and global protein distributions in neurons
Abstract
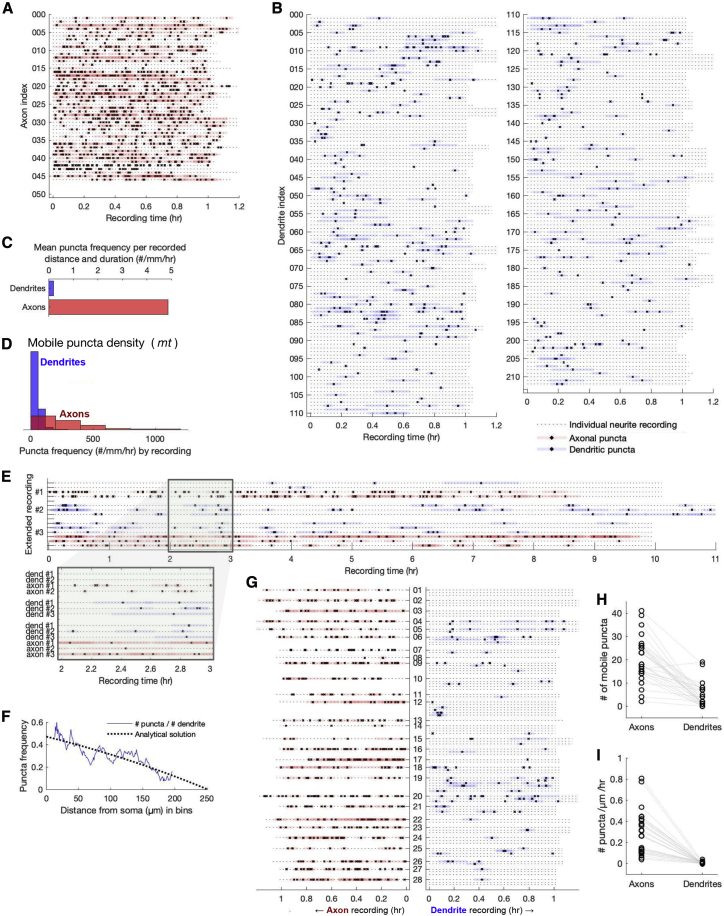
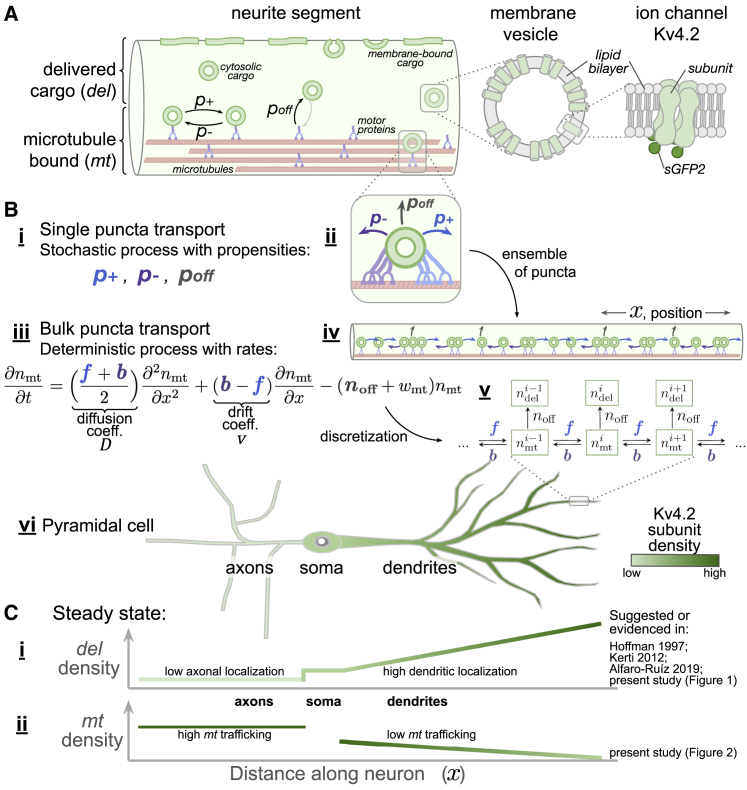
Neural function depends on continual synthesis and targeted trafficking of intracellular components, including ion channel proteins. Many kinds of ion channels are trafficked over long distances to specific cellular compartments. This raises the question of whether cargo is directed with high specificity during transit or whether cargo is distributed widely and sequestered at specific sites. We addressed this question by experimentally measuring transport and expression densities of Kv4.2, a voltage-gated transient potassium channel that exhibits a specific dendritic expression that increases with distance from the soma and little or no functional expression in axons. In over 500 h of quantitative live imaging, we found substantially higher densities of actively transported Kv4.2 subunits in axons as opposed to dendrites. This paradoxical relationship between functional expression and traffic density supports a model-commonly known as the sushi belt model-in which trafficking specificity is relatively low and active sequestration occurs in compartments where cargo is expressed. In further support of this model, we find that kinetics of active transport differs qualitatively between axons and dendrites, with axons exhibiting strong superdiffusivity, whereas dendritic transport resembles a weakly directed random walk, promoting mixing and opportunity for sequestration. Finally, we use our data to constrain a compartmental reaction-diffusion model that can recapitulate the known Kv4.2 density profile. Together, our results show how nontrivial expression patterns can be maintained over long distances with a relatively simple trafficking mechanism and how the hallmarks of a global trafficking mechanism can be revealed in the kinetics and density of cargo.
Copyright © 2021 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures





Similar articles
-
Specific sorting and post-Golgi trafficking of dendritic potassium channels in living neurons.J Biol Chem. 2014 Apr 11;289(15):10566-10581. doi: 10.1074/jbc.M113.534495. Epub 2014 Feb 25. J Biol Chem. 2014. PMID: 24569993 Free PMC article.
-
Dendritic localization and a cis-acting dendritic targeting element of Kv4.2 mRNA.BMB Rep. 2010 Oct;43(10):677-82. doi: 10.5483/BMBRep.2010.43.10.677. BMB Rep. 2010. PMID: 21034530
-
Mossy fibre contact triggers the targeting of Kv4.2 potassium channels to dendrites and synapses in developing cerebellar granule neurons.J Neurochem. 2004 May;89(4):897-907. doi: 10.1111/j.1471-4159.2004.02368.x. J Neurochem. 2004. PMID: 15140189
-
Diverse roles for auxiliary subunits in phosphorylation-dependent regulation of mammalian brain voltage-gated potassium channels.Pflugers Arch. 2011 Nov;462(5):631-43. doi: 10.1007/s00424-011-1004-8. Epub 2011 Aug 6. Pflugers Arch. 2011. PMID: 21822597 Free PMC article. Review.
-
Polarized trafficking: the palmitoylation cycle distributes cytoplasmic proteins to distinct neuronal compartments.Curr Opin Cell Biol. 2018 Feb;50:64-71. doi: 10.1016/j.ceb.2018.02.004. Epub 2018 Feb 21. Curr Opin Cell Biol. 2018. PMID: 29475137 Review.
Cited by
-
Microtubule networks in zebrafish hair cells facilitate presynapse transport and fusion during development.Elife. 2025 Jul 23;13:RP98119. doi: 10.7554/eLife.98119. Elife. 2025. PMID: 40698602 Free PMC article.
-
Computational insights into mRNA and protein dynamics underlying synaptic plasticity rules.Mol Cell Neurosci. 2023 Jun;125:103846. doi: 10.1016/j.mcn.2023.103846. Epub 2023 Mar 22. Mol Cell Neurosci. 2023. PMID: 36963534 Free PMC article. Review.
-
mRNA Trafficking in the Nervous System: A Key Mechanism of the Involvement of Activity-Regulated Cytoskeleton-Associated Protein (Arc) in Synaptic Plasticity.Neural Plast. 2021 Sep 23;2021:3468795. doi: 10.1155/2021/3468795. eCollection 2021. Neural Plast. 2021. PMID: 34603440 Free PMC article. Review.
-
Transport between im/mobile fractions shapes the speed and profile of cargo distribution in neurons.Biophys Rep (N Y). 2022 Oct 20;2(4):100082. doi: 10.1016/j.bpr.2022.100082. eCollection 2022 Dec 14. Biophys Rep (N Y). 2022. PMID: 36425667 Free PMC article.
-
Alzheimer's disease/dementia-associated brain pathology in aging DPP6-KO mice.Neurobiol Dis. 2022 Nov;174:105887. doi: 10.1016/j.nbd.2022.105887. Epub 2022 Oct 6. Neurobiol Dis. 2022. PMID: 36209950 Free PMC article.
References
-
- Kapitein L.C., Hoogenraad C.C. Which way to go? Cytoskeletal organization and polarized transport in neurons. Mol. Cell. Neurosci. 2011;46:9–20. - PubMed
-
- Shibata R., Misonou H., Trimmer J.S. A fundamental role for KChIPs in determining the molecular properties and trafficking of Kv4.2 potassium channels. J. Biol. Chem. 2003;278:36445–36454. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
