

The Origin of Niches and Species in the Bacterial World
- PMID: 33815348
- PMCID: PMC8010147
- DOI: 10.3389/fmicb.2021.657986
The Origin of Niches and Species in the Bacterial World
Abstract
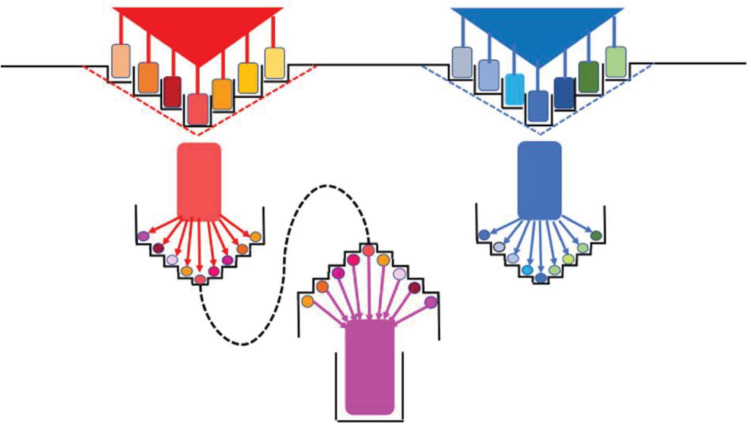
Niches are spaces for the biological units of selection, from cells to complex communities. In a broad sense, "species" are biological units of individuation. Niches do not exist without individual organisms, and every organism has a niche. We use "niche" in the Hutchinsonian sense as an abstraction of a multidimensional environmental space characterized by a variety of conditions, both biotic and abiotic, whose quantitative ranges determine the positive or negative growth rates of the microbial individual, typically a species, but also parts of the communities of species contained in this space. Microbial organisms ("species") constantly diversify, and such diversification (radiation) depends on the possibility of opening up unexploited or insufficiently exploited niches. Niche exploitation frequently implies "niche construction," as the colonized niche evolves with time, giving rise to new potential subniches, thereby influencing the selection of a series of new variants in the progeny. The evolution of niches and organisms is the result of reciprocal interacting processes that form a single unified process. Centrifugal microbial diversification expands the limits of the species' niches while a centripetal or cohesive process occurs simultaneously, mediated by horizontal gene transfers and recombinatorial events, condensing all of the information recovered during the diversifying specialization into "novel organisms" (possible future species), thereby creating a more complex niche, where the selfishness of the new organism(s) establishes a "homeostatic power" limiting the niche's variation. Once the niche's full carrying capacity has been reached, reproductive isolation occurs, as no foreign organisms can outcompete the established population/community, thereby facilitating speciation. In the case of individualization-speciation of the microbiota, its contribution to the animal' gut structure is a type of "niche construction," the result of crosstalk between the niche (host) and microorganism(s). Lastly, there is a parallelism between the hierarchy of niches and that of microbial individuals. The increasing anthropogenic effects on the biosphere (such as globalization) might reduce the diversity of niches and bacterial individuals, with the potential emergence of highly transmissible multispecialists (which are eventually deleterious) resulting from the homogenization of the microbiosphere, a possibility that should be explored and prevented.
Keywords: bacterial niches; bacterial species; evolution; nichification; speciation.
Copyright © 2021 Baquero, Coque, Galán and Martinez.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures

References
-
- Aghajani Delavar M., Wang J. (2020). Modeling combined effects of temperature and structure on competition and growth of multispecies biofilms in microbioreactors. Ind. Eng. Chem. Res. 59 16122–16135. 10.1021/acs.iecr.0c03102 - DOI
-
- Alcaraz L. D., Olmedo G., Bonilla G., Cerritos R., Hernández G., Cruz A., et al. (2008). The genome of Bacillus coahuilensis reveals adaptations essential for survival in the relic of an ancient marine environment. Proc. Natl. Acad. Sci. U.S.A. 105 5803–5808. 10.1073/pnas.0800981105 - DOI - PMC - PubMed
-
- Baquero F. (1994). “The ecological code,” in Descifrar la Vida: Ensayos de Historia de la Biología, eds Casadesús J., Ruiz Berraquero F. (Seville: Seville University Press; ), 355–365.
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources