

Subunit Vaccines Using TLR Triagonist Combination Adjuvants Provide Protection Against Coxiella burnetii While Minimizing Reactogenic Responses
- PMID: 33815413
- PMCID: PMC8010241
- DOI: 10.3389/fimmu.2021.653092
Subunit Vaccines Using TLR Triagonist Combination Adjuvants Provide Protection Against Coxiella burnetii While Minimizing Reactogenic Responses
Abstract
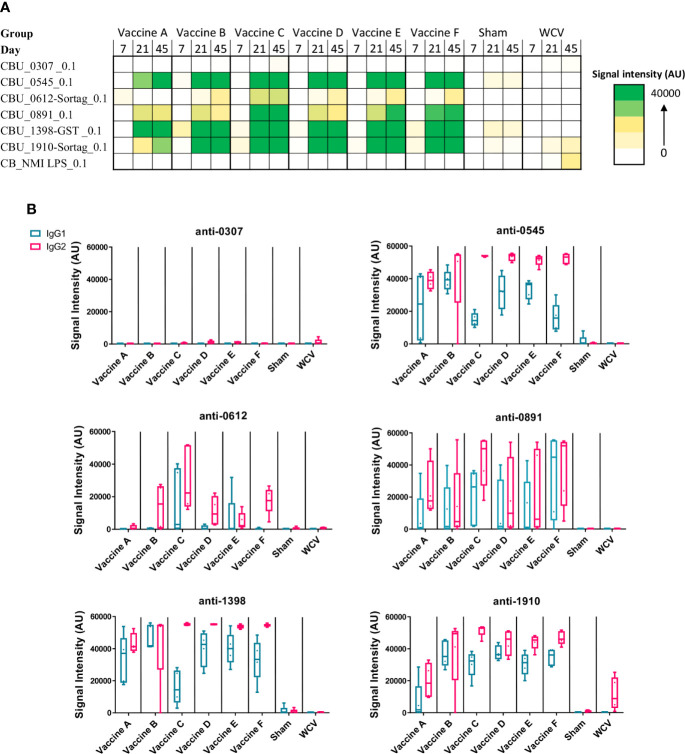
Q fever is caused by the obligate intracellular bacterium, Coxiella burnetii, a designated potential agent of bioterrorism because of its route of transmission, resistance to disinfectants, and low infectious dose. The only vaccine licensed for human use is Q-VAX® (Seqirus, licensed in Australia), a formalin-inactivated whole-cell vaccine, which produces severe local and systemic reactogenic responses in previously sensitized individuals. Accordingly, the U.S. Food and Drug Administration and other regulatory bodies around the world, have been reluctant to approve Q-VAX for widespread use. To obviate these adverse reactions, we prepared recombinant protein subunit vaccine candidates containing purified CBU1910, CBU0307, CBU0545, CBU0612, CBU0891, and CBU1398 proteins and TLR triagonist adjuvants. TLR triagonist adjuvants combine different TLR agonists to enhance immune responses to vaccine antigens. We tested both the protective efficacy and reactogenicity of our vaccine candidates in Hartley guinea pigs using intratracheal infection with live C. burnetii. While all of our candidates showed varying degrees of protection during challenge, local reactogenic responses were significantly reduced for one of our vaccine candidates when compared with a formalin-inactivated whole-cell vaccine. Our findings show that subunit vaccines combined with novel TLR triagonist adjuvants can generate protective immunity to C. burnetii infection while reducing reactogenic responses.
Keywords: Coxiella burnetii; TLR agonist; guinea pig model; hypersensitivity; triagonist; vaccine.
Copyright © 2021 Fratzke, Jan, Felgner, Liang, Nakajima, Jasinskas, Manna, Nihesh, Maiti, Albin, Esser-Kahn, Davies, Samuel, Felgner and Gregory.
Conflict of interest statement
PF, DD, JF, LL, SJ, RN, and AJ own shares in Nanommune Inc. Nanommune does not sell the arrays described in this paper, nor funded any part of the work described herein. Neither Nanommune or its shareholders are likely to benefit from the results described in this publication. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Tuning Subunit Vaccines with Novel TLR Triagonist Adjuvants to Generate Protective Immune Responses against Coxiella burnetii.J Immunol. 2020 Feb 1;204(3):611-621. doi: 10.4049/jimmunol.1900991. Epub 2019 Dec 23. J Immunol. 2020. PMID: 31871024
-
Multivalent vaccines demonstrate immunogenicity and protect against Coxiella burnetii aerosol challenge.Front Immunol. 2023 Jul 18;14:1192821. doi: 10.3389/fimmu.2023.1192821. eCollection 2023. Front Immunol. 2023. PMID: 37533862 Free PMC article.
-
Coxiella burnetii Whole Cell Vaccine Produces a Th1 Delayed-Type Hypersensitivity Response in a Novel Sensitized Mouse Model.Front Immunol. 2021 Sep 20;12:754712. doi: 10.3389/fimmu.2021.754712. eCollection 2021. Front Immunol. 2021. PMID: 34616410 Free PMC article.
-
Immunogenicity and Reactogenicity in Q Fever Vaccine Development.Front Immunol. 2022 May 26;13:886810. doi: 10.3389/fimmu.2022.886810. eCollection 2022. Front Immunol. 2022. PMID: 35693783 Free PMC article. Review.
-
Components of protective immunity.Adv Exp Med Biol. 2012;984:91-104. doi: 10.1007/978-94-007-4315-1_5. Adv Exp Med Biol. 2012. PMID: 22711628 Review.
Cited by
-
Preclinical Animal Models for Q Fever Vaccine Development.Front Cell Infect Microbiol. 2022 Feb 10;12:828784. doi: 10.3389/fcimb.2022.828784. eCollection 2022. Front Cell Infect Microbiol. 2022. PMID: 35223553 Free PMC article. Review.
-
Soluble antigens derived from Coxiella burnetii elicit protective immunity in three animal models without inducing hypersensitivity.Cell Rep Med. 2021 Dec 6;2(12):100461. doi: 10.1016/j.xcrm.2021.100461. eCollection 2021 Dec 21. Cell Rep Med. 2021. PMID: 35028605 Free PMC article.
-
Q fever immunology: the quest for a safe and effective vaccine.NPJ Vaccines. 2023 Sep 7;8(1):133. doi: 10.1038/s41541-023-00727-6. NPJ Vaccines. 2023. PMID: 37679410 Free PMC article. Review.
-
Recent Advances on the Innate Immune Response to Coxiella burnetii.Front Cell Infect Microbiol. 2021 Nov 2;11:754455. doi: 10.3389/fcimb.2021.754455. eCollection 2021. Front Cell Infect Microbiol. 2021. PMID: 34796128 Free PMC article. Review.
-
Molecular mechanisms of Coxiella burnetii formalin-fixed cellular vaccine reactogenicity.Infect Immun. 2024 Nov 12;92(11):e0033524. doi: 10.1128/iai.00335-24. Epub 2024 Oct 2. Infect Immun. 2024. PMID: 39356158 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources