

When the Ends Justify the Means: Regulation of Telomere Addition at Double-Strand Breaks in Yeast
- PMID: 33816507
- PMCID: PMC8012806
- DOI: 10.3389/fcell.2021.655377
When the Ends Justify the Means: Regulation of Telomere Addition at Double-Strand Breaks in Yeast
Abstract
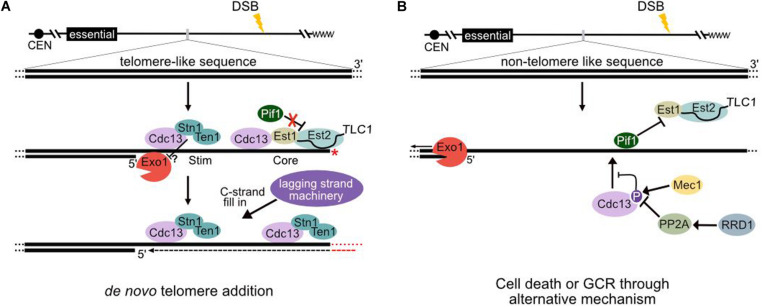
Telomeres, repetitive sequences located at the ends of most eukaryotic chromosomes, provide a mechanism to replenish terminal sequences lost during DNA replication, limit nucleolytic resection, and protect chromosome ends from engaging in double-strand break (DSB) repair. The ribonucleoprotein telomerase contains an RNA subunit that serves as the template for the synthesis of telomeric DNA. While telomere elongation is typically primed by a 3' overhang at existing chromosome ends, telomerase can act upon internal non-telomeric sequences. Such de novo telomere addition can be programmed (for example, during chromosome fragmentation in ciliated protozoa) or can occur spontaneously in response to a chromosome break. Telomerase action at a DSB can interfere with conservative mechanisms of DNA repair and results in loss of distal sequences but may prevent additional nucleolytic resection and/or chromosome rearrangement through formation of a functional telomere (termed "chromosome healing"). Here, we review studies of spontaneous and induced DSBs in the yeast Saccharomyces cerevisiae that shed light on mechanisms that negatively regulate de novo telomere addition, in particular how the cell prevents telomerase action at DSBs while facilitating elongation of critically short telomeres. Much of our understanding comes from the use of perfect artificial telomeric tracts to "seed" de novo telomere addition. However, endogenous sequences that are enriched in thymine and guanine nucleotides on one strand (TG-rich) but do not perfectly match the telomere consensus sequence can also stimulate unusually high frequencies of telomere formation following a DSB. These observations suggest that some internal sites may fully or partially escape mechanisms that normally negatively regulate de novo telomere addition.
Keywords: DNA repair; Pif1; de novo telomere addition; telomerase; telomere.
Copyright © 2021 Hoerr, Ngo and Friedman.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Interaction of yeast Rad51 and Rad52 relieves Rad52-mediated inhibition of de novo telomere addition.PLoS Genet. 2020 Feb 3;16(2):e1008608. doi: 10.1371/journal.pgen.1008608. eCollection 2020 Feb. PLoS Genet. 2020. PMID: 32012161 Free PMC article.
-
Regulation of telomere addition at DNA double-strand breaks.Chromosoma. 2013 Jun;122(3):159-73. doi: 10.1007/s00412-013-0404-2. Epub 2013 Mar 17. Chromosoma. 2013. PMID: 23504035 Review.
-
Chromosome healing, telomere capture and mechanisms of radiation-induced chromosome breakage.Int J Radiat Biol. 1998 Jan;73(1):1-13. doi: 10.1080/095530098142653. Int J Radiat Biol. 1998. PMID: 9464472 Review.
-
Endogenous Hot Spots of De Novo Telomere Addition in the Yeast Genome Contain Proximal Enhancers That Bind Cdc13.Mol Cell Biol. 2016 May 31;36(12):1750-63. doi: 10.1128/MCB.00095-16. Print 2016 Jun 15. Mol Cell Biol. 2016. PMID: 27044869 Free PMC article.
-
Chromosome healing by de novo telomere addition in Saccharomyces cerevisiae.Mol Microbiol. 2006 Mar;59(5):1357-68. doi: 10.1111/j.1365-2958.2006.05026.x. Mol Microbiol. 2006. PMID: 16468981 Review.
Cited by
-
The ubiquitin protease Ubp10 suppresses the formation of translocations at Cdc13 binding sites.bioRxiv [Preprint]. 2025 Jun 28:2025.06.27.661970. doi: 10.1101/2025.06.27.661970. bioRxiv. 2025. PMID: 40667018 Free PMC article. Preprint.
-
A proto-telomere is elongated by telomerase in a shelterin-dependent manner in quiescent fission yeast cells.Nucleic Acids Res. 2022 Nov 11;50(20):11682-11695. doi: 10.1093/nar/gkac986. Nucleic Acids Res. 2022. PMID: 36330920 Free PMC article.
-
A comprehensive map of hotspots of de novo telomere addition in Saccharomyces cerevisiae.Genetics. 2023 May 26;224(2):iyad076. doi: 10.1093/genetics/iyad076. Genetics. 2023. PMID: 37119805 Free PMC article.
-
Hotspot of de novo telomere addition stabilizes linear amplicons in yeast grown in sulfate-limiting conditions.Genetics. 2023 May 26;224(2):iyad010. doi: 10.1093/genetics/iyad010. Genetics. 2023. PMID: 36702776 Free PMC article.
-
A comprehensive map of hotspots of de novo telomere addition in Saccharomyces cerevisiae.bioRxiv [Preprint]. 2023 Mar 23:2023.03.20.533556. doi: 10.1101/2023.03.20.533556. bioRxiv. 2023. Update in: Genetics. 2023 May 26;224(2):iyad076. doi: 10.1093/genetics/iyad076. PMID: 36993206 Free PMC article. Updated. Preprint.
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous