

In vivo N-Terminomics Highlights Novel Functions of ADAMTS2 and ADAMTS14 in Skin Collagen Matrix Building
- PMID: 33816558
- PMCID: PMC8017238
- DOI: 10.3389/fmolb.2021.643178
In vivo N-Terminomics Highlights Novel Functions of ADAMTS2 and ADAMTS14 in Skin Collagen Matrix Building
Abstract
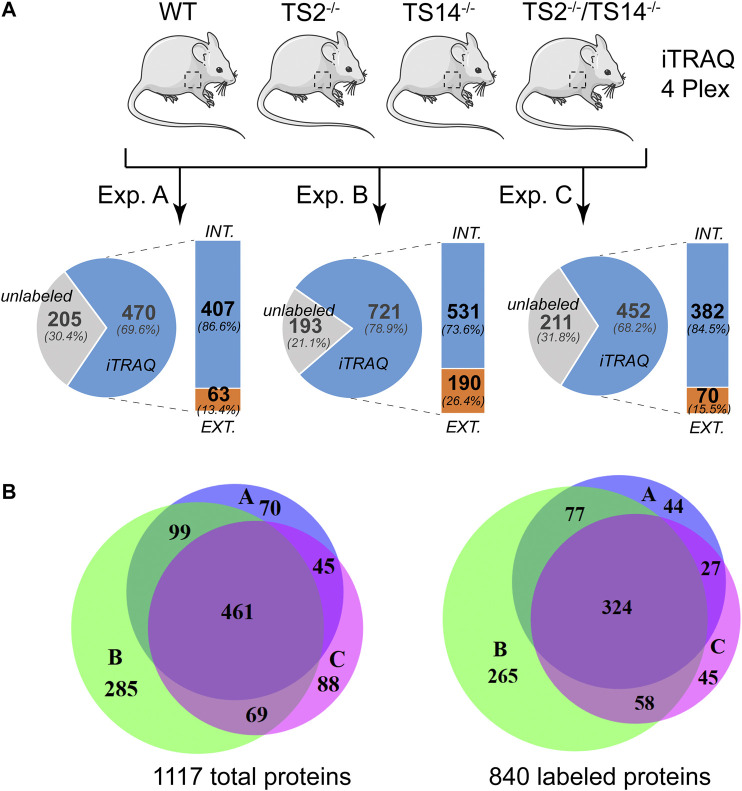
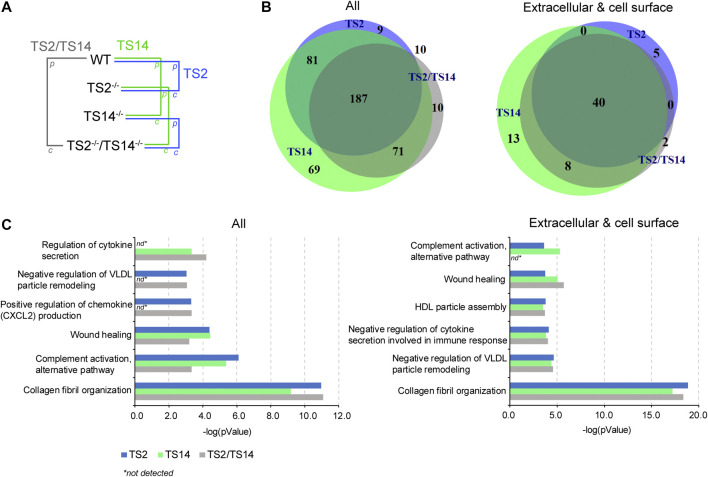
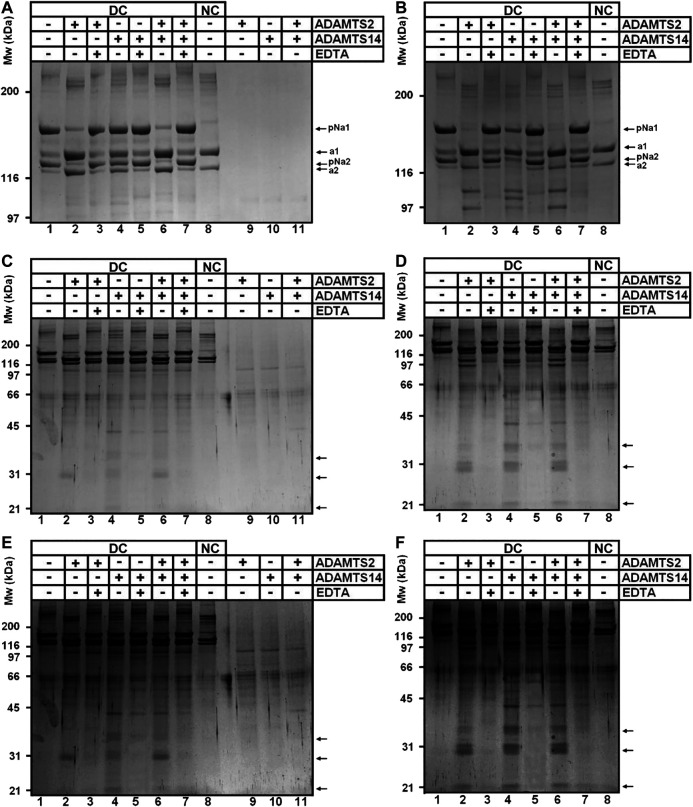
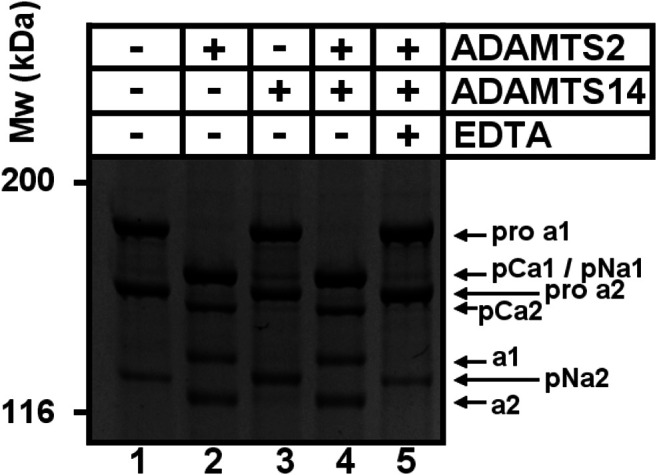
A disintegrin and metalloproteinase with thrombospondin type I motif (ADAMTS)2 and ADAMTS14 were originally known for their ability to cleave the aminopropeptides of fibrillar collagens. Previous work using N-terminomic approach (N-TAILS) in vitro led to the identification of new substrates, including some molecules involved in TGF-β signaling. Here, N-TAILS was used to investigate the substrates of these two enzymes in vivo, by comparing the N-terminomes of the skin of wild type mice, mice deficient in ADAMTS2, in ADAMTS14 and in both ADAMTS2 and ADAMTS14. This study identified 68 potential extracellular and cell surface proteins, with the majority of them being cleaved by both enzymes. These analyses comfort their role in collagen matrix organization and suggest their implication in inflammatory processes. Regarding fibrillar collagen, this study demonstrates that both ADAMTS2 and ADAMTS14 are involved in the processing of the aminopropeptide of alpha1 and alpha2 type V collagen. It also revealed the existence of several cleavage sites in the Col1 domain and in the C-propeptide of type I collagens. In addition to collagens and other extracellular proteins, two major components of the cell cytoskeleton, actin and vimentin, were also identified as potential substrates. The latter data were confirmed in vitro using purified enzymes and could potentially indicate other functions for ADAMTS2 and 14. This original investigation of mouse skin degradomes by N-terminomic highlights the essential role of ADAMTS2 and ADAMTS14 in collagen matrix synthesis and turnover, and gives clues to better understand their functions in skin pathophysiology. Data are available via ProteomeXchange with identifier PXD022179.
Keywords: ADAMTS; Ehlers-Danlos Syndrome (EDS); N-Terminomics; TAILS; collagen; degradomics.
Copyright © 2021 Leduc, Dupont, Joannes, Monseur, Baiwir, Mazzucchelli, Deroanne, Colige and Bekhouche.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures



Similar articles
-
Regulation of procollagen amino-propeptide processing during mouse embryogenesis by specialization of homologous ADAMTS proteases: insights on collagen biosynthesis and dermatosparaxis.Development. 2006 Apr;133(8):1587-96. doi: 10.1242/dev.02308. Development. 2006. PMID: 16556917
-
Spontaneous atopic dermatitis due to immune dysregulation in mice lacking Adamts2 and 14.Matrix Biol. 2018 Sep;70:140-157. doi: 10.1016/j.matbio.2018.04.002. Epub 2018 Apr 9. Matrix Biol. 2018. PMID: 29649548
-
Determination of the substrate repertoire of ADAMTS2, 3, and 14 significantly broadens their functions and identifies extracellular matrix organization and TGF-β signaling as primary targets.FASEB J. 2016 May;30(5):1741-56. doi: 10.1096/fj.15-279869. Epub 2016 Jan 6. FASEB J. 2016. PMID: 26740262
-
The procollagen N-proteinases ADAMTS2, 3 and 14 in pathophysiology.Matrix Biol. 2015 May-Jul;44-46:46-53. doi: 10.1016/j.matbio.2015.04.001. Epub 2015 Apr 8. Matrix Biol. 2015. PMID: 25863161 Review.
-
[The Ehlers-Danlos syndrome: the extracellular matrix scaffold in question].Med Sci (Paris). 2003 Apr;19(4):443-52. doi: 10.1051/medsci/2003194443. Med Sci (Paris). 2003. PMID: 12836217 Review. French.
Cited by
-
Artificial neural network identified the significant genes to distinguish Idiopathic pulmonary fibrosis.Sci Rep. 2023 Jan 21;13(1):1225. doi: 10.1038/s41598-023-28536-w. Sci Rep. 2023. PMID: 36681777 Free PMC article.
-
The Pathogenesis of Cardiac Fibrosis: A Review of Recent Progress.Int J Mol Sci. 2022 Feb 27;23(5):2617. doi: 10.3390/ijms23052617. Int J Mol Sci. 2022. PMID: 35269759 Free PMC article. Review.
-
The Mechanism and Role of ADAMTS Protein Family in Osteoarthritis.Biomolecules. 2022 Jul 8;12(7):959. doi: 10.3390/biom12070959. Biomolecules. 2022. PMID: 35883515 Free PMC article. Review.
-
The ADAMTS2 metalloproteinase inhibits tumor growth by regulating the innate immune system.Cancer Cell Int. 2025 Jun 23;25(1):229. doi: 10.1186/s12935-025-03880-1. Cancer Cell Int. 2025. PMID: 40551102 Free PMC article.
-
Phylogenetic inference of the emergence of sequence modules and protein-protein interactions in the ADAMTS-TSL family.PLoS Comput Biol. 2023 Aug 31;19(8):e1011404. doi: 10.1371/journal.pcbi.1011404. eCollection 2023 Aug. PLoS Comput Biol. 2023. PMID: 37651409 Free PMC article.
References
-
- Beighton P., De Paepe A., Steinmann B., Tsipouras P., Wenstrup R. J. (1998). Ehlers-danlos syndromes: revised nosology, villefranche, 1997. Ehlers-danlos national foundation (United States) and ehlers-danlos support group (United Kindom). Am. J. Med. Genet. 77, 31–37. 10.1002/(sici)1096-8628(19980428)77:1<31::aid-ajmg8>3.0.co;2-o - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous