

Epithelia-Sensory Neuron Cross Talk Underlies Cholestatic Itch Induced by Lysophosphatidylcholine
- PMID: 33819485
- PMCID: PMC9093619
- DOI: 10.1053/j.gastro.2021.03.049
Epithelia-Sensory Neuron Cross Talk Underlies Cholestatic Itch Induced by Lysophosphatidylcholine
Abstract
Background & aims: Limited understanding of pruritus mechanisms in cholestatic liver diseases hinders development of antipruritic treatments. Previous studies implicated lysophosphatidic acid (LPA) as a potential mediator of cholestatic pruritus.
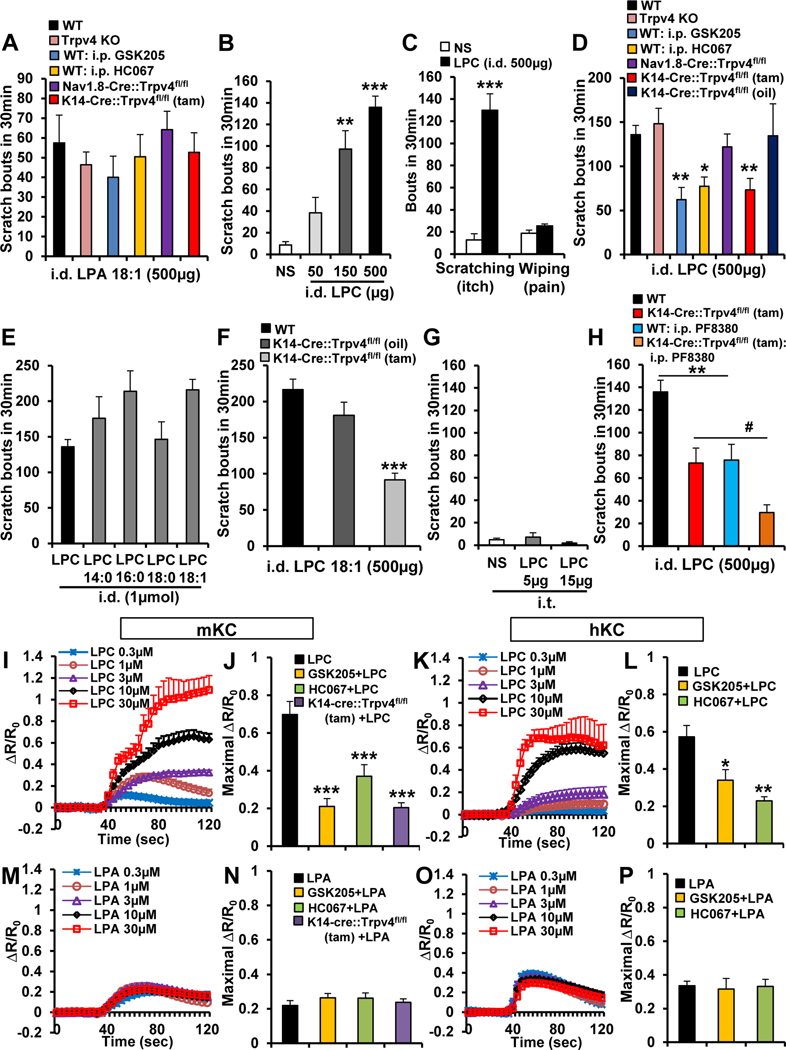
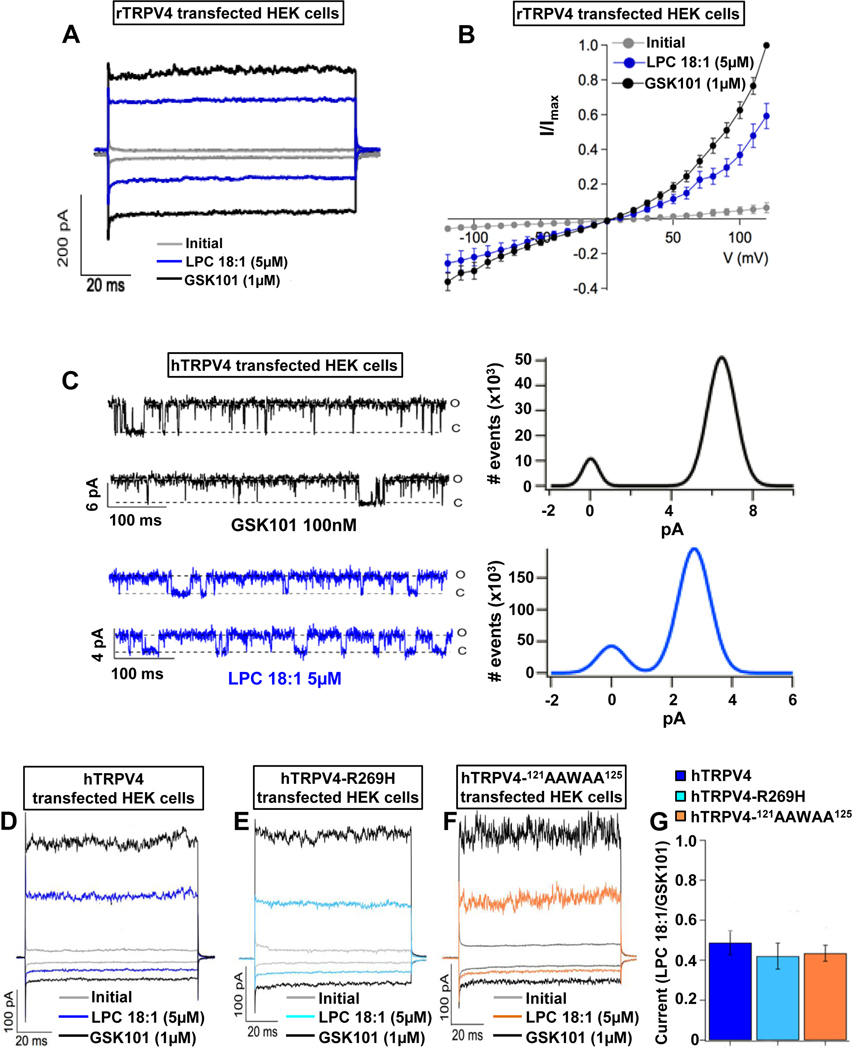
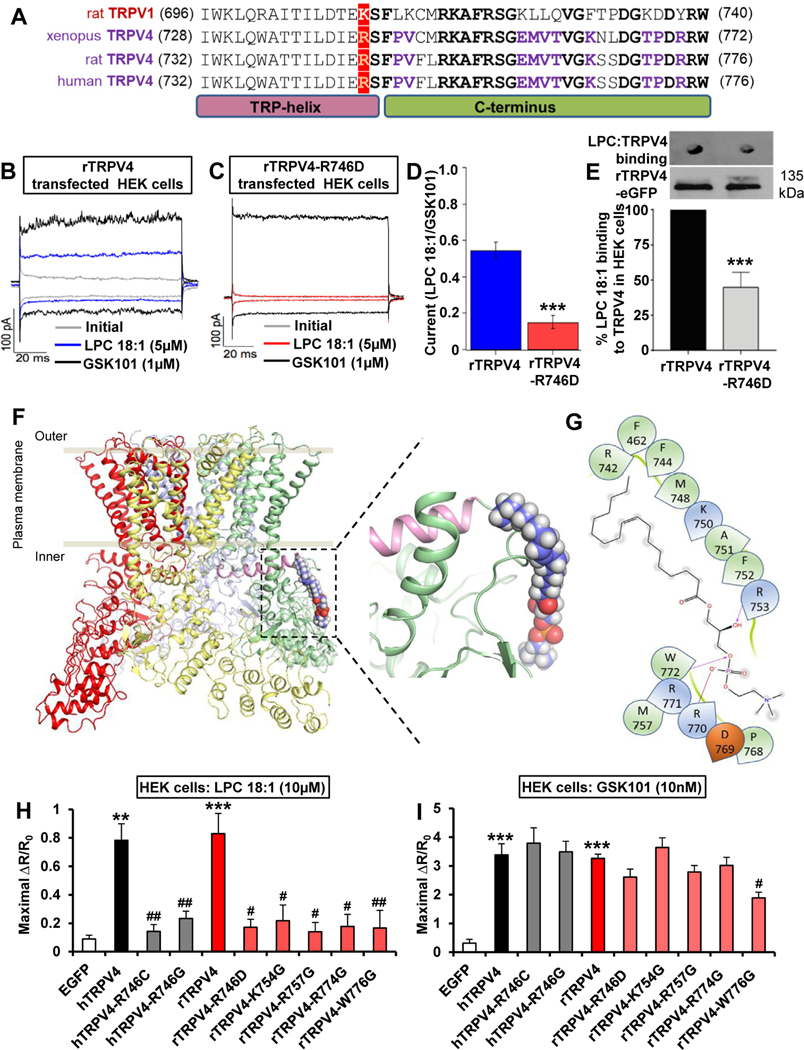
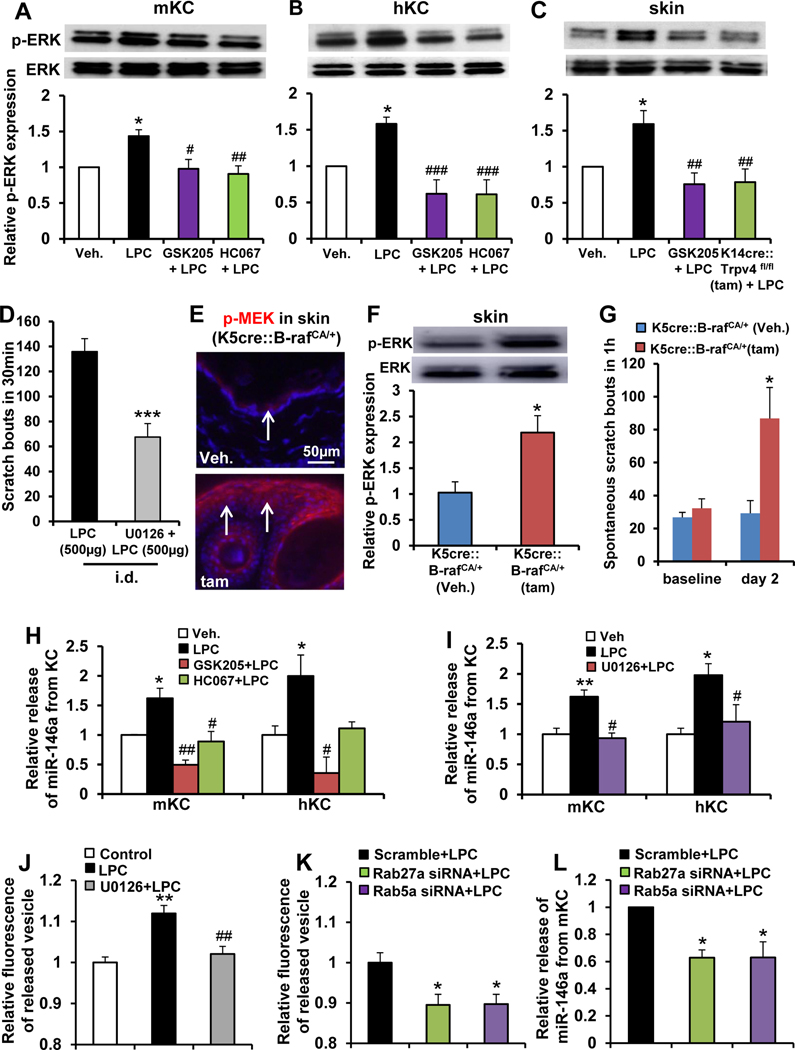
Methods: Pruritogenicity of lysophosphatidylcholine (LPC), LPA's precursor, was examined in naïve mice, cholestatic mice, and nonhuman primates. LPC's pruritogenicity involving keratinocyte TRPV4 was studied using genetic and pharmacologic approaches, cultured keratinocytes, ion channel physiology, and structural computational modeling. Activation of pruriceptor sensory neurons by microRNA-146a (miR-146a), secreted from keratinocytes, was identified by in vitro and ex vivo Ca2+ imaging assays. Sera from patients with primary biliary cholangitis were used for measuring the levels of LPC and miR-146a.
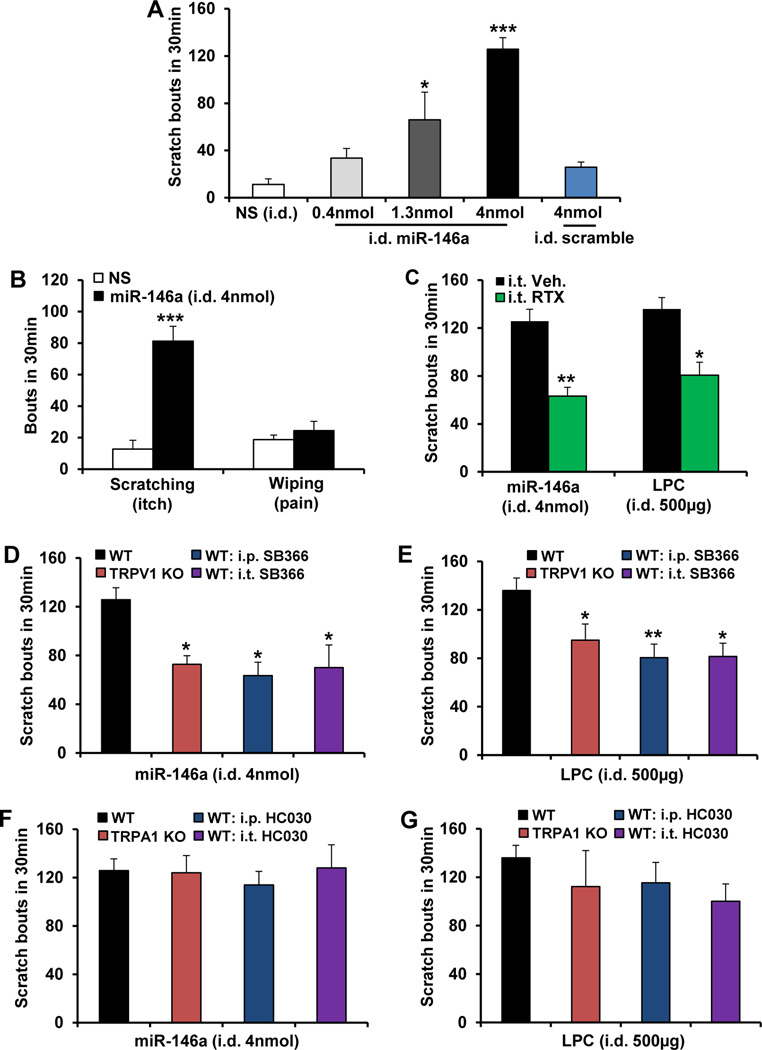
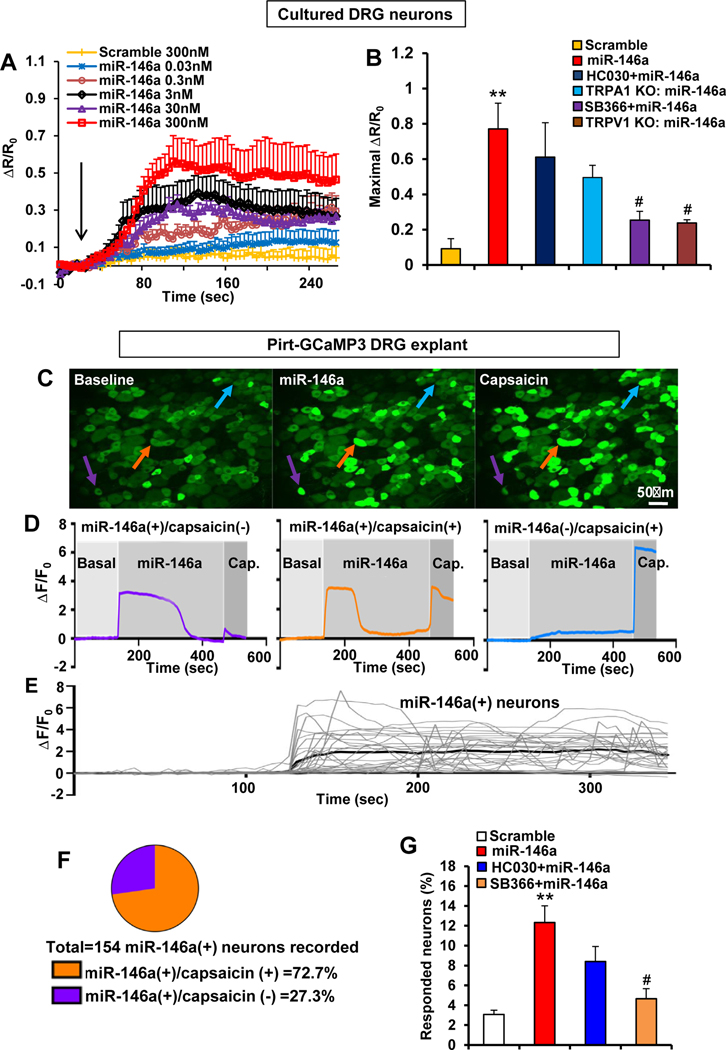
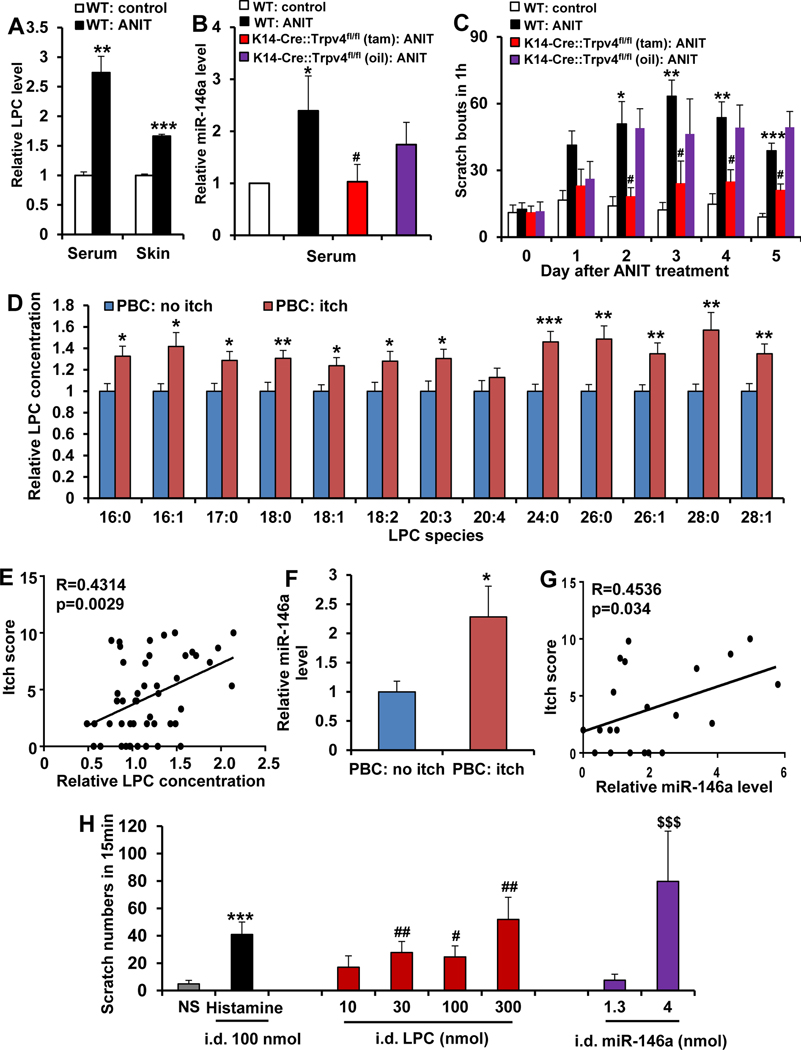
Results: LPC was robustly pruritic in mice. TRPV4 in skin keratinocytes was essential for LPC-induced itch and itch in mice with cholestasis. Three-dimensional structural modeling, site-directed mutagenesis, and channel function analysis suggested a TRPV4 C-terminal motif for LPC binding and channel activation. In keratinocytes, TRPV4 activation by LPC induced extracellular release of miR-146a, which activated TRPV1+ sensory neurons to cause itch. LPC and miR-146a levels were both elevated in sera of patients with primary biliary cholangitis with itch and correlated with itch intensity. Moreover, LPC and miR-146a were also increased in sera of cholestatic mice and elicited itch in nonhuman primates.
Conclusions: We identified LPC as a novel cholestatic pruritogen that induces itch through epithelia-sensory neuron cross talk, whereby it directly activates skin keratinocyte TRPV4, which rapidly releases miR-146a to activate skin-innervating TRPV1+ pruriceptor sensory neurons. Our findings support the new concept of the skin, as a sensory organ, playing a critical role in cholestatic itch, beyond liver, peripheral sensory neurons, and central neural pathways supporting pruriception.
Keywords: Cholestatic Itch; Keratinocyte TRPV4; Lysophosphatidylcholine; TRPV1 Pruriceptor; miR-146a.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflicts of interest
The authors disclose no conflicts.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
