

Blood leukocytes recapitulate diabetogenic peptide-MHC-II complexes displayed in the pancreatic islets
- PMID: 33822842
- PMCID: PMC8034384
- DOI: 10.1084/jem.20202530
Blood leukocytes recapitulate diabetogenic peptide-MHC-II complexes displayed in the pancreatic islets
Abstract
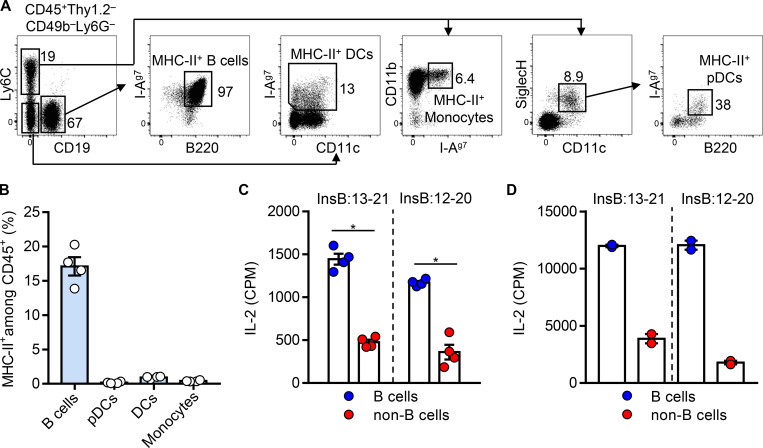
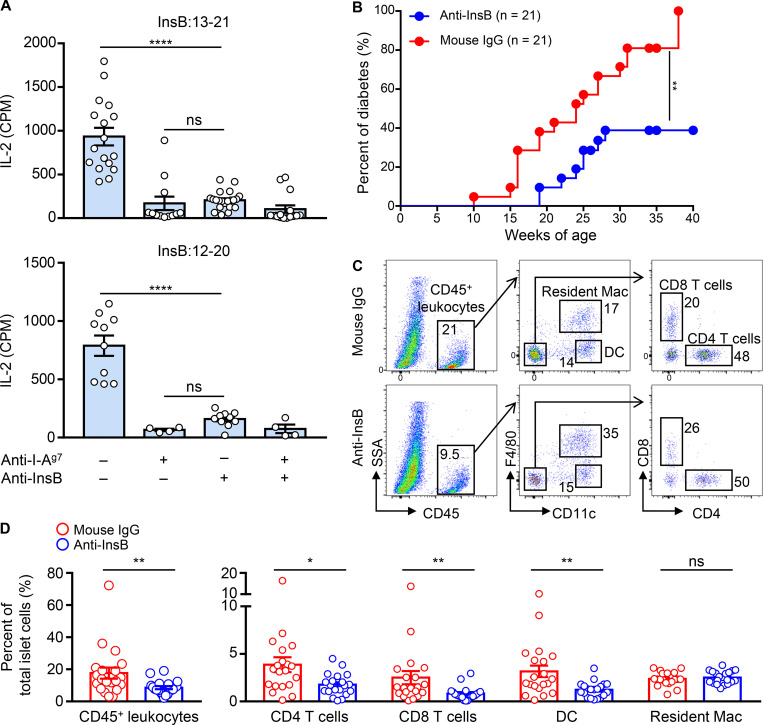
Assessing the self-peptides presented by susceptible major histocompatibility complex (MHC) molecules is crucial for evaluating the pathogenesis and therapeutics of tissue-specific autoimmune diseases. However, direct examination of such MHC-bound peptides displayed in the target organ remains largely impractical. Here, we demonstrate that the blood leukocytes from the nonobese diabetic (NOD) mice presented peptide epitopes to autoreactive CD4 T cells. These peptides were bound to the autoimmune class II MHC molecule (MHC-II) I-Ag7 and originated from insulin B-chain and C-peptide. The presentation required a glucose challenge, which stimulated the release of the insulin peptides from the pancreatic islets. The circulating leukocytes, especially the B cells, promptly captured and presented these peptides. Mass spectrometry analysis of the leukocyte MHC-II peptidome revealed a series of β cell-derived peptides, with identical sequences to those previously identified in the islet MHC-II peptidome. Thus, the blood leukocyte peptidome echoes that found in islets and serves to identify immunogenic peptides in an otherwise inaccessible tissue.
© 2021 Vomund et al.
Conflict of interest statement
Disclosures: The authors declare no competing interests exist.
Figures


References
-
- Babon, J.A.B., DeNicola M.E., Blodgett D.M., Crèvecoeur I., Buttrick T.S., Maehr R., Bottino R., Naji A., Kaddis J., Elyaman W., et al. . 2016. Analysis of self-antigen specificity of islet-infiltrating T cells from human donors with type 1 diabetes. Nat. Med. 22:1482–1487. 10.1038/nm.4203 - DOI - PMC - PubMed
-
- Carrero, J.A., McCarthy D.P., Ferris S.T., Wan X., Hu H., Zinselmeyer B.H., Vomund A.N., and Unanue E.R.. 2017. Resident macrophages of pancreatic islets have a seminal role in the initiation of autoimmune diabetes of NOD mice. Proc. Natl. Acad. Sci. USA. 114:E10418–E10427. 10.1073/pnas.1713543114 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
