

Kir4.1/Kir5.1 channels possess strong intrinsic inward rectification determined by a voltage-dependent K+-flux gating mechanism
- PMID: 33822868
- PMCID: PMC8025212
- DOI: 10.1085/jgp.201912540
Kir4.1/Kir5.1 channels possess strong intrinsic inward rectification determined by a voltage-dependent K+-flux gating mechanism
Abstract
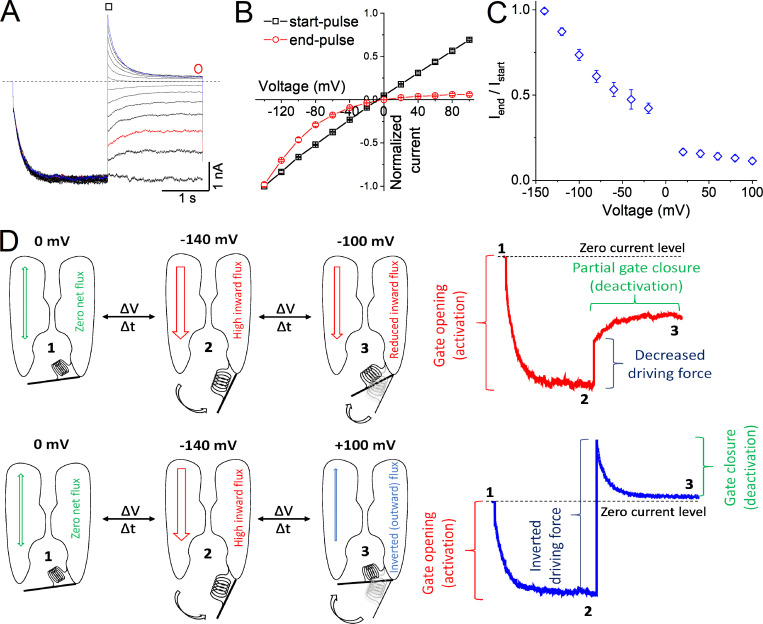
Inwardly rectifying potassium (Kir) channels are broadly expressed in both excitable and nonexcitable tissues, where they contribute to a wide variety of cellular functions. Numerous studies have established that rectification of Kir channels is not an inherent property of the channel protein itself, but rather reflects strong voltage dependence of channel block by intracellular cations, such as polyamines and Mg2+. Here, we identify a previously unknown mechanism of inward rectification in Kir4.1/Kir5.1 channels in the absence of these endogenous blockers. This novel intrinsic rectification originates from the voltage-dependent behavior of Kir4.1/Kir5.1, which is generated by the flux of potassium ions through the channel pore; the inward K+-flux induces the opening of the gate, whereas the outward flux is unable to maintain the gate open. This gating mechanism powered by the K+-flux is convergent with the gating of PIP2 because, at a saturating concentration, PIP2 greatly reduces the inward rectification. Our findings provide evidence of the coexistence of two rectification mechanisms in Kir4.1/Kir5.1 channels: the classical inward rectification induced by blocking cations and an intrinsic voltage-dependent mechanism generated by the K+-flux gating.
© 2021 Marmolejo-Murillo et al.
Figures






Comment in
-
Unconventional voltage sensing in an inwardly rectifying potassium channel.J Gen Physiol. 2021 Jun 7;153(6):e202112929. doi: 10.1085/jgp.202112929. Epub 2021 May 6. J Gen Physiol. 2021. PMID: 33956072 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
