

Nearest-neighbour transition-state analysis for nucleic acid kinetics
- PMID: 33823552
- PMCID: PMC8096236
- DOI: 10.1093/nar/gkab205
Nearest-neighbour transition-state analysis for nucleic acid kinetics
Abstract
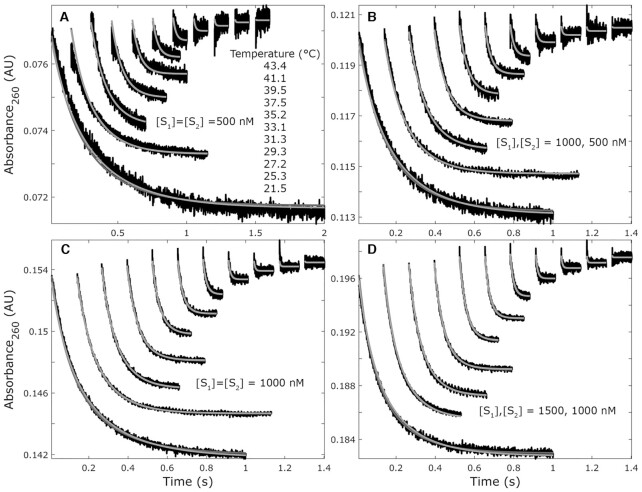
We used stopped-flow to monitor hypochromicity for 43 oligonucleotide duplexes to study nucleic acid kinetics and extract transition-state parameters for association and dissociation. Reactions were performed in 1.0 M NaCl (for literature comparisons) and 2.2 mM MgCl2 (PCR conditions). Dissociation kinetics depended on sequence, increased exponentially with temperature, and transition-state parameters inversely correlated to thermodynamic parameters (r = -0.99). Association had no consistent enthalpic component, varied little with temperature or sequence, and poorly correlated to thermodynamic parameters (r = 0.28). Average association rates decreased 78% in MgCl2 compared to NaCl while dissociation was relatively insensitive to ionic conditions. A nearest-neighbour kinetic model for dissociation predicted rate constants within 3-fold of literature values (n = 11). However, a nearest-neighbour model for association appeared overparameterized and inadequate for predictions. Kinetic predictions were used to simulate published high-speed (<1 min) melting analysis and extreme (<2 min) PCR experiments. Melting simulations predicted apparent melting temperatures increase on average 2.4°C when temperature ramp rates increased from 0.1 to 32°C/s, compared to 2.8°C reported in the literature. PCR simulations revealed that denaturation kinetics are dependent on the thermocycling profile. Simulations overestimated annealing efficiencies at shorter annealing times and suggested that polymerase interactions contribute to primer-template complex stability at extension temperatures.
© The Author(s) 2021. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures





References
-
- Borer P.N., Dengler B., Tinoco I., Uhlenbeck O.C.. Stability of ribonucleic acid double-stranded helices. J. Mol. Biol. 1974; 86:843–853. - PubMed
-
- Myrick J.T., Pryor R.J., Palais R.A., Ison S.J., Sanford L., Dwight Z.L., Huuskonen J.J., Sundberg S.O., Wittwer C.T.. Integrated extreme real-time PCR and high-speed melting analysis in 52 to 87 seconds. Clin. Chem. 2019; 65:263–271. - PubMed
-
- Rejali N.A., Zuiter A.M., Quackenbush J.F., Wittwer C.T.. Reverse transcriptase kinetics for one-step RT-PCR. Anal. Biochem. 2020; 601:113768. - PubMed
-
- Wetmur J.G. Hybridization and renaturation kinetics of nucleic acids. Annu. Rev. Biophys. Bioeng. 1976; 5:337–361. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
