

The genetic feature and virulence determinant of highly virulent community-associated MRSA ST338-SCCmec Vb in China
- PMID: 33823746
- PMCID: PMC8183566
- DOI: 10.1080/22221751.2021.1914516
The genetic feature and virulence determinant of highly virulent community-associated MRSA ST338-SCCmec Vb in China
Abstract
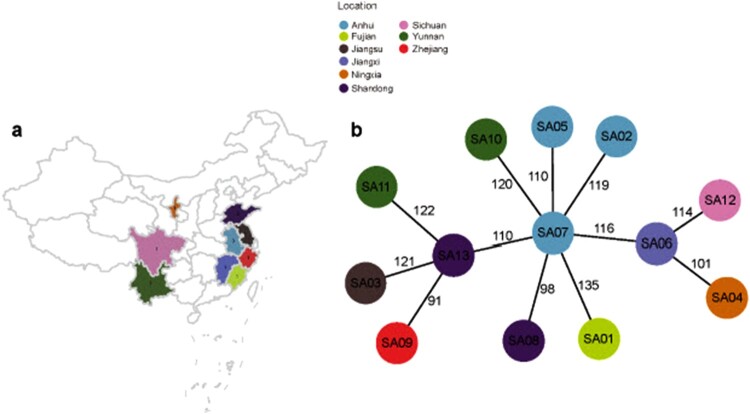
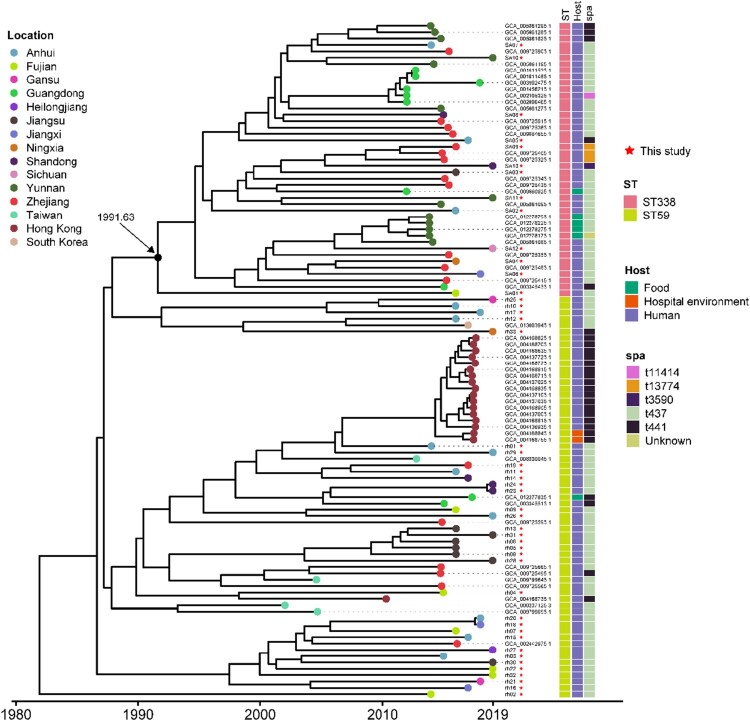
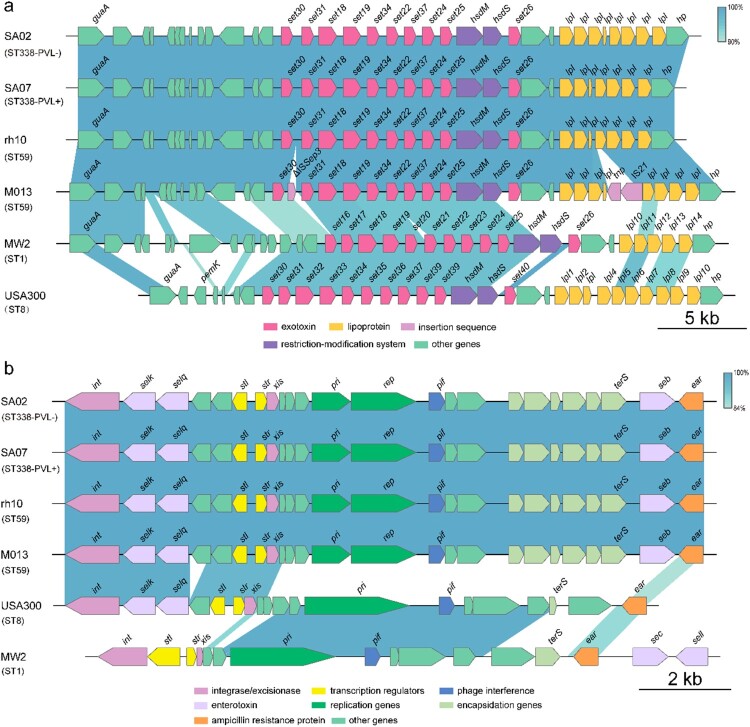
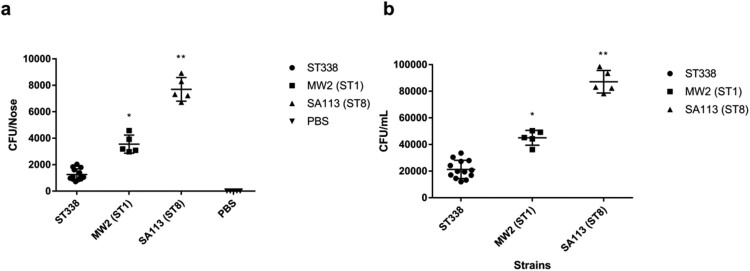
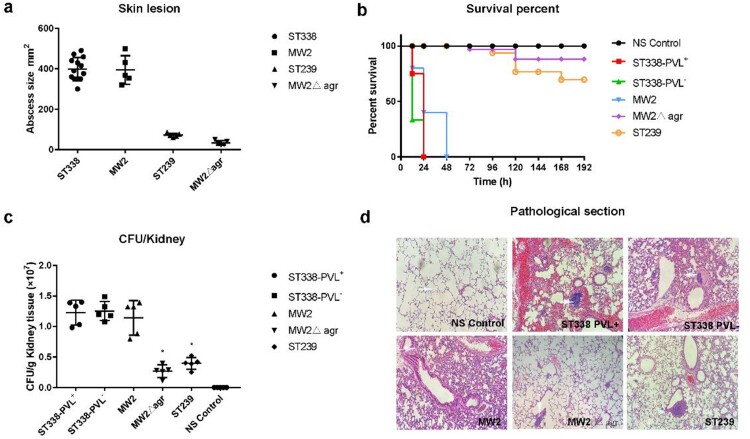
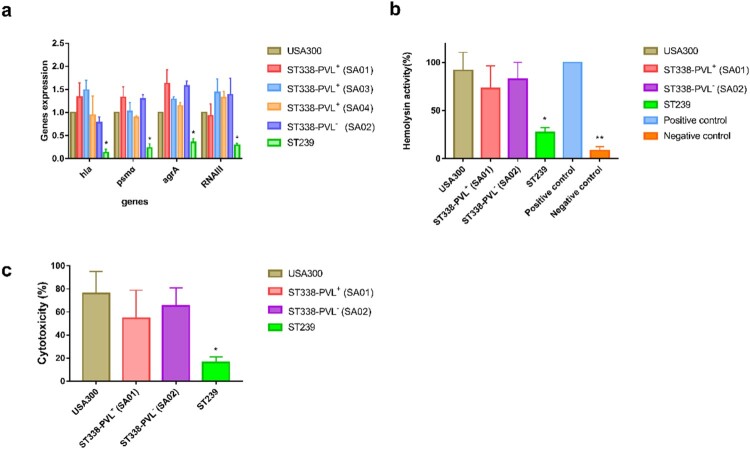
ST59 is the predominant pathotype of community-associated methicillin-resistant Staphylococcus aureus (CA-MRSA) in China. As a variant of ST59, there is relatively little known about the detailed information of ST338. To address this issue, here, we described thirteen ST338 CA-MRSA strains isolated from severe bloodstream infection cases, and focused on their epidemiology, genetic features and virulence potential. Phylogenetic analysis showed the earliest isolated strain of this study is likely a predecessor of recent ST338 lineage (after year of 2014). Furthermore, the phylogenetic reconstruction and time estimation suggested that ST338 evolved from ST59 in 1991. Notably, the carrying patten of virulence factors of all ST338 strains were similar, and the genomic islands νSaα, νSaγ and SaPI and the core virulence factors like hla and psm were detected in ST338 isolates. However, all ST338 isolates lacked some adhesion factors such as clfA, clfB, eap, cna and icaD. Additionally, among these ST338 strains, one PVL-negative ST338 isolate was detected. Experiment on mice nose and human alveolar epithelial cell showed that the nasal colonization ability of ST338 was weaker than that of CA-MRSA MW2. In a mouse bloodstream infection model and skin infection model, PVL+ and PVL- strains had the similar virulence, which was dependent on upregulation of toxin genes rather than the presence of mobile genetic elements such as ΦSa2 carrying PVL. Our findings provide important insight into the epidemiology and pathogenicity of the novel and highly virulent ST338-SCCmec Vb clone.
Keywords: CA-MRSA; Panton–Valentine leucocidin; ST338; bloodstream infection; core-virulence factors.
Conflict of interest statement
No potential conflict of interest was reported by the author(s).
Figures

References
-
- Tacconelli E, Tumbarello M, Cauda R.. Staphylococcus aureus infections. N Engl J Med. 1998;339:2026–2027. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous