

Limits of temperature adaptation and thermopreferendum
- PMID: 33823918
- PMCID: PMC8025563
- DOI: 10.1186/s13578-021-00574-9
Limits of temperature adaptation and thermopreferendum
Abstract
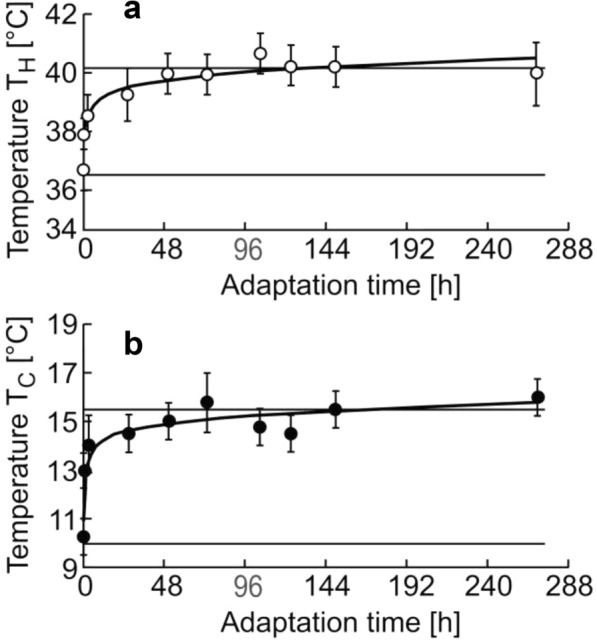
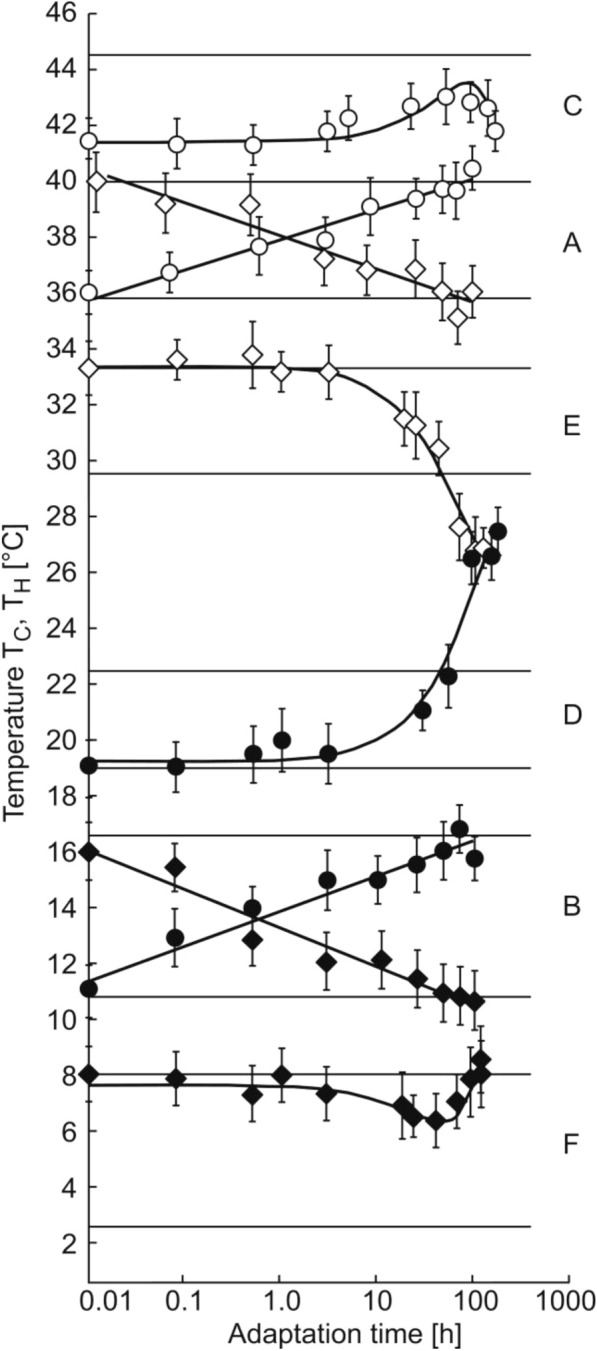
Background: Managing the limits of temperature adaptation is relevant both in medicine and in biotechnology. There are numerous scattered publications on the identification of the temperature limits of existence for various organisms and using different methods. Dmitry Petrovich Kharakoz gave a general explanation for many of these experimental results. The hypothesis implied that each cycle of synaptic exocytosis includes reversible phase transitions of lipids of the presynaptic membrane due to the entry and subsequent removal of calcium ions from the synaptic terminal. The correspondence of the times of phase transitions has previously been experimentally shown on isolated lipids in vitro. In order to test the hypothesis of D.P. Kharakoz in vivo, we investigated the influence of the temperature of long-term acclimatization on the temperature of heat and cold shock, as well as on the kinetics of temperature adaptation in zebrafish. Testing the hypothesis included a comparison of our experimental results with the results of other authors obtained on various models from invertebrates to humans.
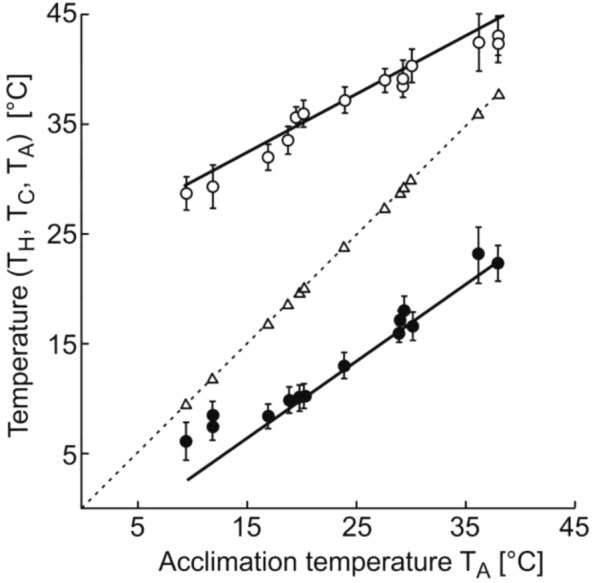
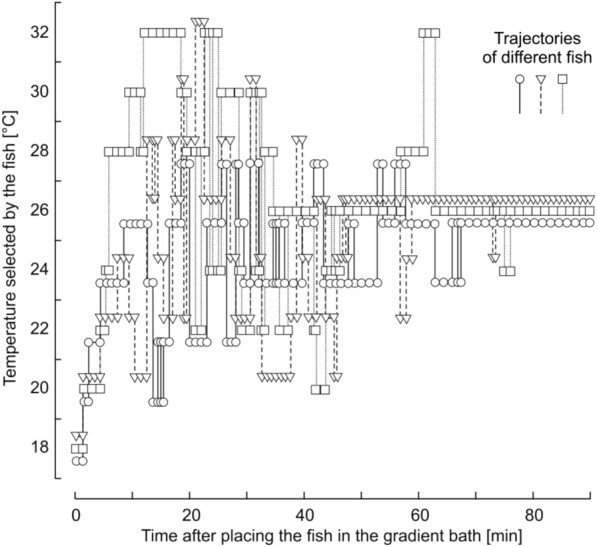
Results: The viability polygon for Danio rerio was determined by the minimum temperature of cold shock (about 6 °C), maximum temperature of heat shock (about 43 °C), and thermopreferendum temperature (about 27 °C). The ratio of the temperature range of cold shock to the temperature range of heat shock was about 1.3. These parameters obtained for Danio rerio describe with good accuracy those for the planarian Girardia tigrina, the ground squirrel Sermophilus undulatus, and for Homo sapiens.
Conclusions: The experimental values of the temperatures of cold shock and heat shock and the temperature of the thermal preferendum correspond to the temperatures of phase transitions of the lipid-protein composition of the synaptic membrane between the liquid and solid states. The viability range for zebrafish coincides with the temperature range, over which enzymes function effectively and also coincides with the viability polygons for the vast majority of organisms. The boundaries of the viability polygon are characteristic biological constants. The viability polygon of a particular organism is determined not only by the genome, but also by the physicochemical properties of lipids that make up the membrane structures of synaptic endings. The limits of temperature adaptation of any biological species are determined by the temperature range of the functioning of its nervous system.
Keywords: Adaptation; Cold shock; Heat shock; Lipids; Temperature; Thermopreferendum; Viability polygon.
Conflict of interest statement
Authors have no conflict of interests. This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Figures


References
-
- Kharakoz DP. Phase transition in lipids and the problem of homoiothermia. Biofizika. 2000;45(3):569–572. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
