

Input-specific regulation of glutamatergic synaptic transmission in the medial prefrontal cortex by mGlu2/mGlu4 receptor heterodimers
- PMID: 33824180
- PMCID: PMC8279257
- DOI: 10.1126/scisignal.abd2319
Input-specific regulation of glutamatergic synaptic transmission in the medial prefrontal cortex by mGlu2/mGlu4 receptor heterodimers
Abstract
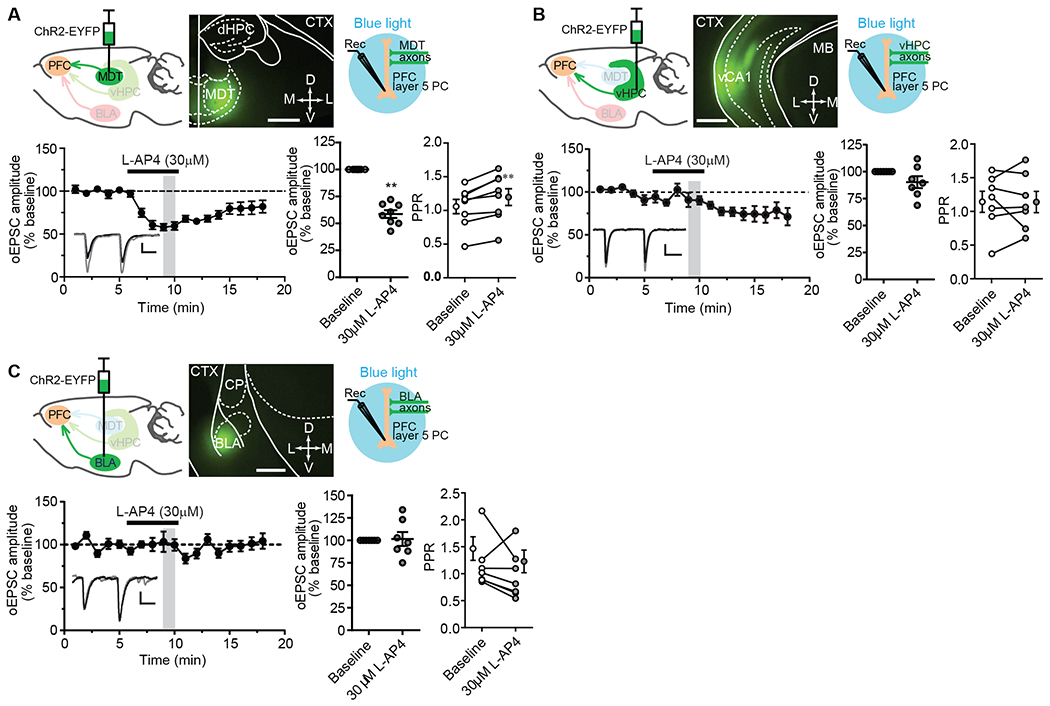
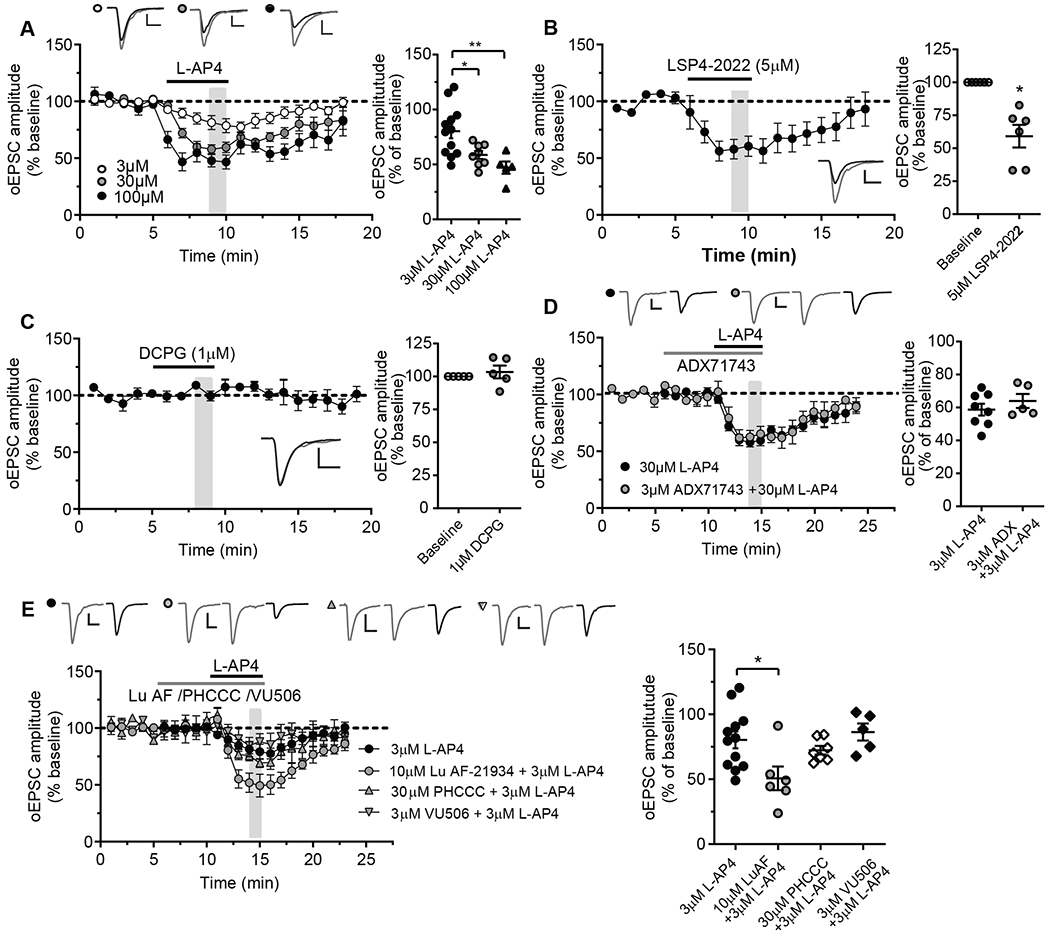
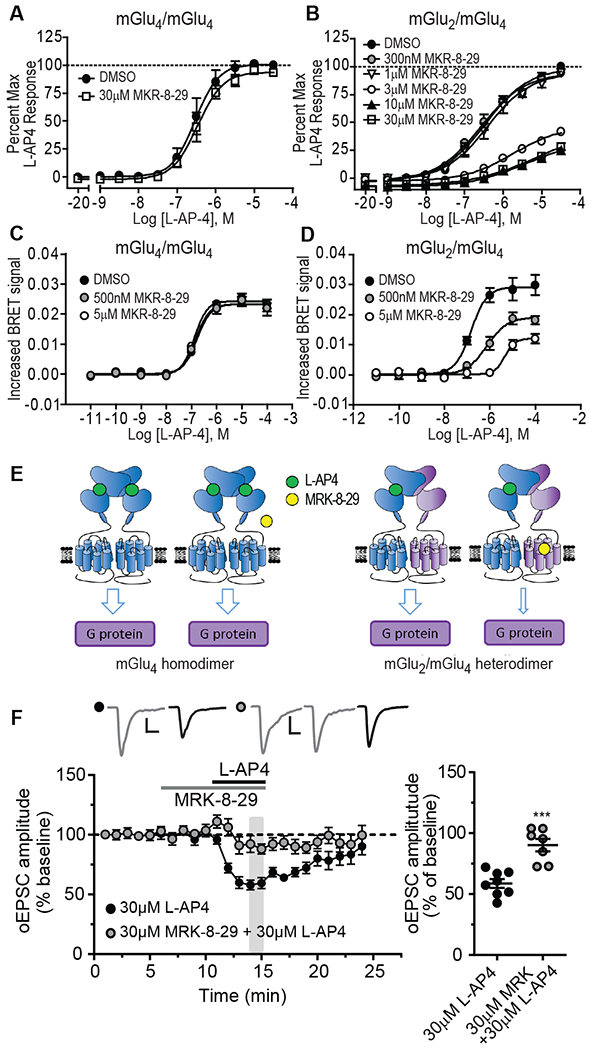
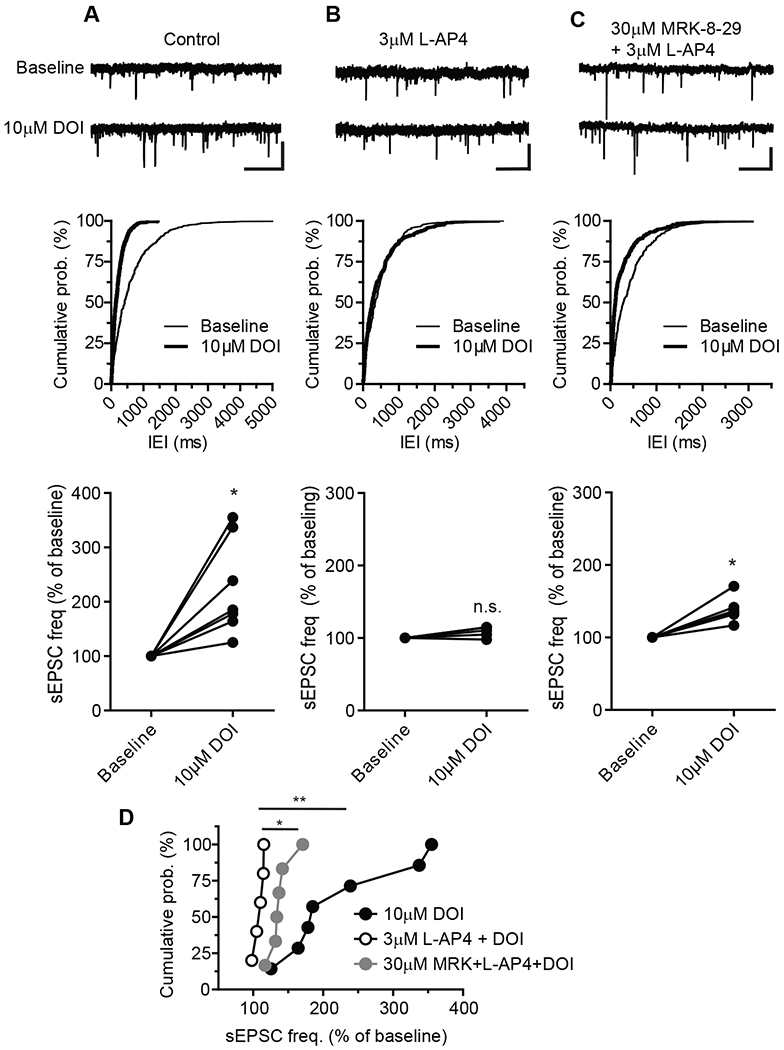
Metabotropic glutamate receptors (mGluRs) are G protein-coupled receptors that regulate various aspects of central nervous system processing in normal physiology and in disease. They are thought to function as disulfide-linked homodimers, but studies have suggested that mGluRs can form functional heterodimers in cell lines. Using selective allosteric ligands, ex vivo brain slice electrophysiology, and optogenetic approaches, we found that two mGluR subtypes-mGluR2 and mGluR4 (or mGlu2 and mGlu4)-exist as functional heterodimers that regulate excitatory transmission in a synapse-specific manner within the rodent medial prefrontal cortex (mPFC). Activation of mGlu2/mGlu4 heterodimers inhibited glutamatergic signaling at thalamo-mPFC synapses but not at hippocampus-mPFC or amygdala-mPFC synapses. These findings raise the possibility that selectively targeting these heterodimers could be a therapeutic strategy for some neurologic and neuropsychiatric disorders involving specific brain circuits.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
References
-
- Nicoletti F, Bockaert J, Collingridge GL, Conn PJ, Ferraguti F, Schoepp DD, Wroblewski JT, Pin JP, Metabotropic glutamate receptors: from the workbench to the bedside. Neuropharmacology 60, 1017–1041 (2011); published online Epub6/2011 (S0028-3908(10)00292-3 [pii];10.1016/j.neuropharm.2010.10.022 [doi]). - DOI - PMC - PubMed
-
- Romano C, Yang WL, O’Malley KL, Metabotropic glutamate receptor 5 is a disulfide-linked dimer. J.Biol.Chem 271, 28612–28616 (1996); published online Epub11/8/1996 ( - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
