

Robustness and plasticity in Drosophila heat avoidance
- PMID: 33824330
- PMCID: PMC8024383
- DOI: 10.1038/s41467-021-22322-w
Robustness and plasticity in Drosophila heat avoidance
Abstract
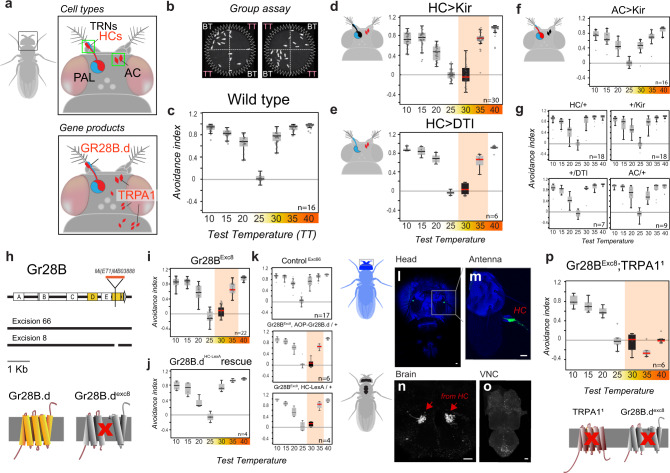
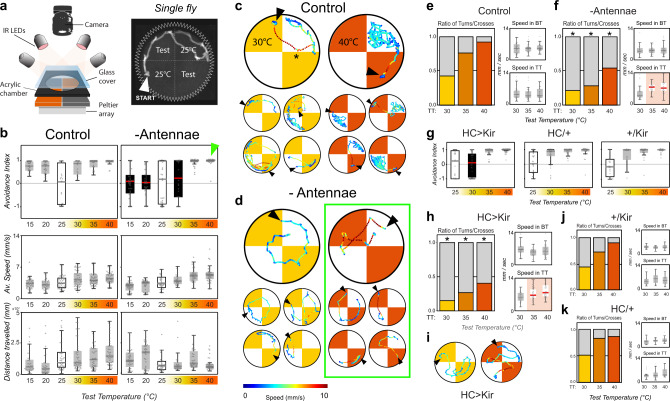
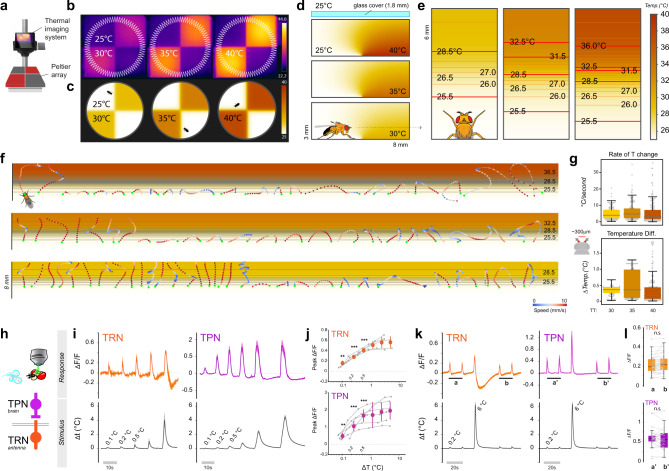
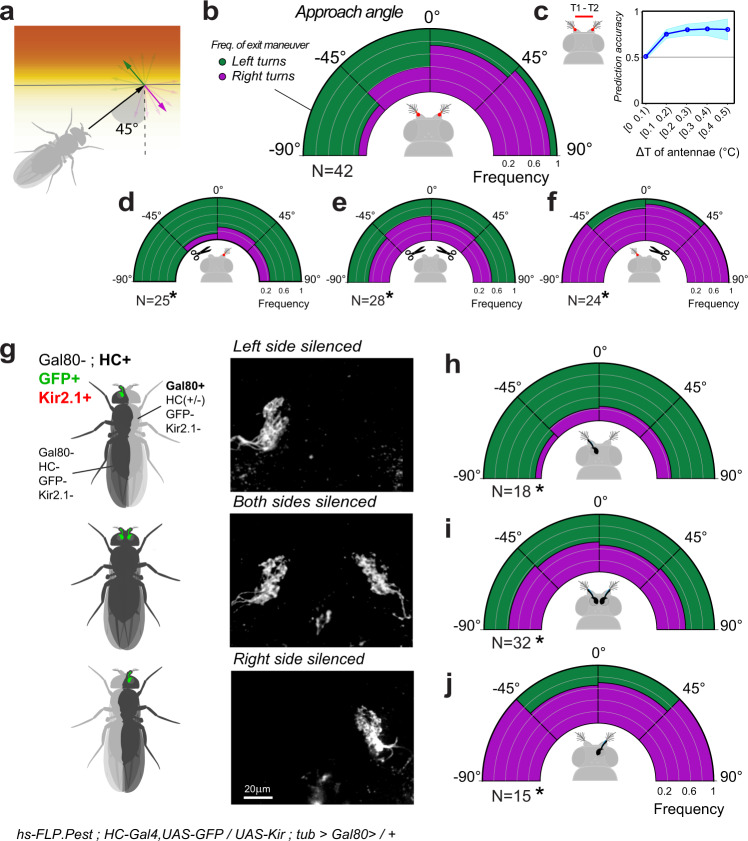
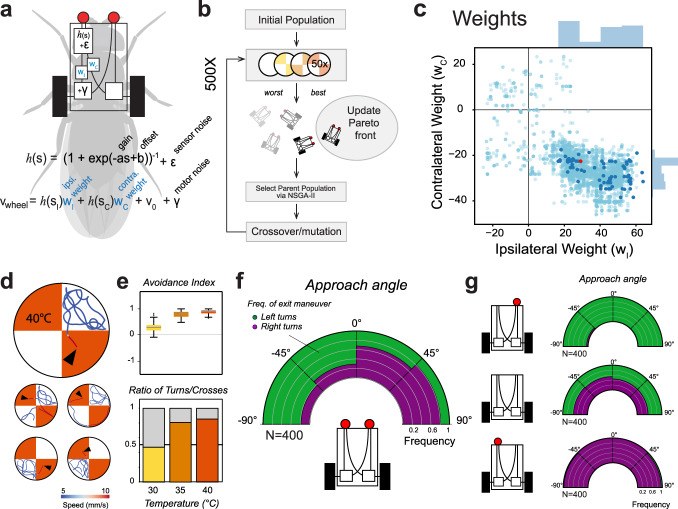
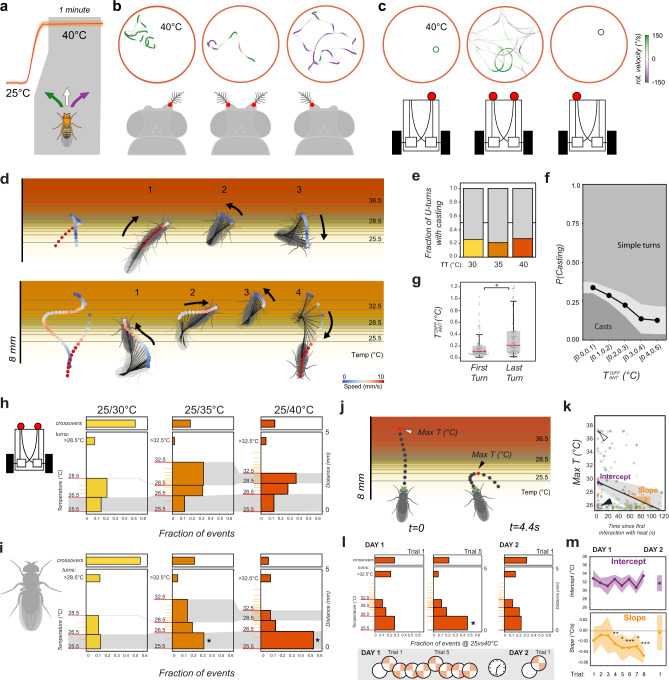
Simple innate behavior is often described as hard-wired and largely inflexible. Here, we show that the avoidance of hot temperature, a simple innate behavior, contains unexpected plasticity in Drosophila. First, we demonstrate that hot receptor neurons of the antenna and their molecular heat sensor, Gr28B.d, are essential for flies to produce escape turns away from heat. High-resolution fly tracking combined with a 3D simulation of the thermal environment shows that, in steep thermal gradients, the direction of escape turns is determined by minute temperature differences between the antennae (0.1°-1 °C). In parallel, live calcium imaging confirms that such small stimuli reliably activate both peripheral thermosensory neurons and central circuits. Next, based on our measurements, we evolve a fly/vehicle model with two symmetrical sensors and motors (a "Braitenberg vehicle") which closely approximates basic fly thermotaxis. Critical differences between real flies and the hard-wired vehicle reveal that fly heat avoidance involves decision-making, relies on rapid learning, and is robust to new conditions, features generally associated with more complex behavior.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Braitenberg, V. Vehicles, Experiments in Synthetic Psychology (MIT Press, 1984).
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
