

The structure of Prp2 bound to RNA and ADP-BeF3- reveals structural features important for RNA unwinding by DEAH-box ATPases
- PMID: 33825710
- PMCID: PMC8025883
- DOI: 10.1107/S2059798321001194
The structure of Prp2 bound to RNA and ADP-BeF3- reveals structural features important for RNA unwinding by DEAH-box ATPases
Abstract
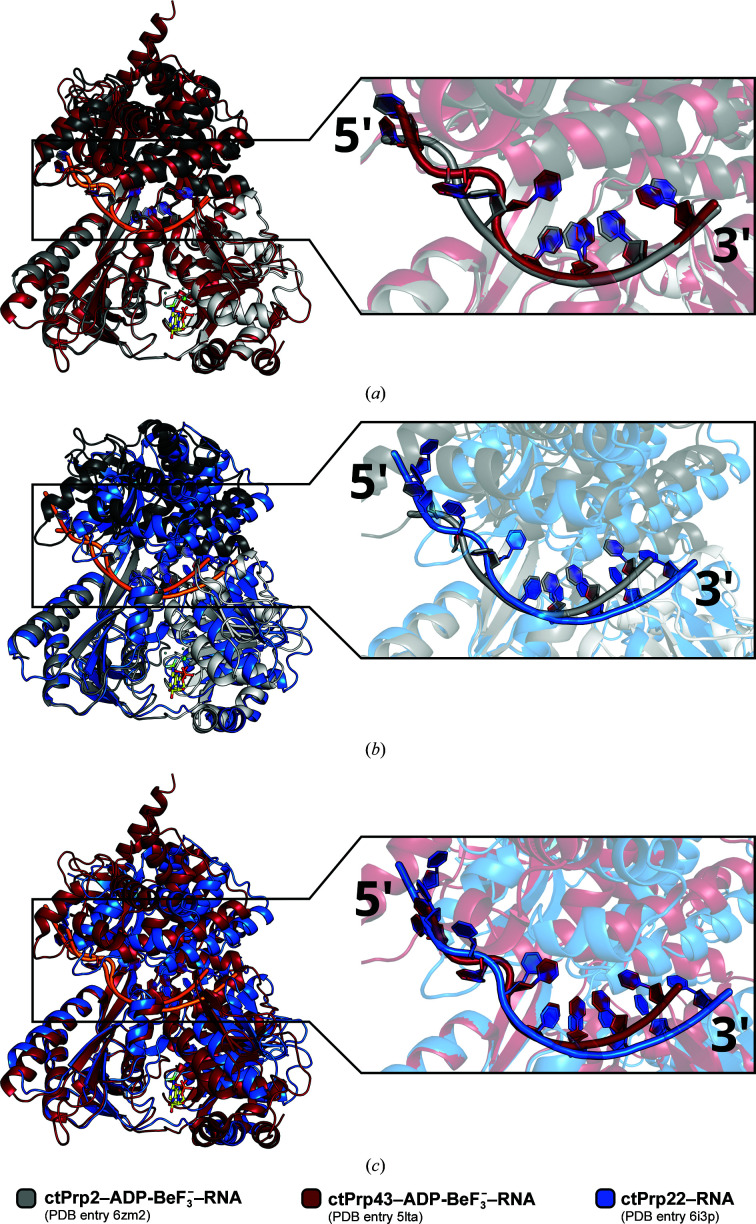
Noncoding intron sequences present in precursor mRNAs need to be removed prior to translation, and they are excised via the spliceosome, a multimegadalton molecular machine composed of numerous protein and RNA components. The DEAH-box ATPase Prp2 plays a crucial role during pre-mRNA splicing as it ensures the catalytic activation of the spliceosome. Despite high structural similarity to other spliceosomal DEAH-box helicases, Prp2 does not seem to function as an RNA helicase, but rather as an RNA-dependent ribonucleoprotein particle-modifying ATPase. Recent crystal structures of the spliceosomal DEAH-box ATPases Prp43 and Prp22, as well as of the related RNA helicase MLE, in complex with RNA have contributed to a better understanding of how RNA binding and processivity might be achieved in this helicase family. In order to shed light onto the divergent manner of function of Prp2, an N-terminally truncated construct of Chaetomium thermophilum Prp2 was crystallized in the presence of ADP-BeF3- and a poly-U12 RNA. The refined structure revealed a virtually identical conformation of the helicase core compared with the ADP-BeF3-- and RNA-bound structure of Prp43, and only a minor shift of the C-terminal domains. However, Prp2 and Prp43 differ in the hook-loop and a loop of the helix-bundle domain, which interacts with the hook-loop and evokes a different RNA conformation immediately after the 3' stack. On replacing these loop residues in Prp43 by the Prp2 sequence, the unwinding activity of Prp43 was abolished. Furthermore, a putative exit tunnel for the γ-phosphate after ATP hydrolysis could be identified in one of the Prp2 structures.
Keywords: DEAH-box ATPases; Prp2; Prp43; RNA helicases; spliceosome.
open access.
Figures





Similar articles
-
Crystal structure of the spliceosomal DEAH-box ATPase Prp2.Acta Crystallogr D Struct Biol. 2018 Jul 1;74(Pt 7):643-654. doi: 10.1107/S2059798318006356. Epub 2018 Jun 8. Acta Crystallogr D Struct Biol. 2018. PMID: 29968674 Free PMC article.
-
Structural basis for RNA translocation by DEAH-box ATPases.Nucleic Acids Res. 2019 May 7;47(8):4349-4362. doi: 10.1093/nar/gkz150. Nucleic Acids Res. 2019. PMID: 30828714 Free PMC article.
-
Structural insights into the mechanism of the DEAH-box RNA helicase Prp43.Elife. 2017 Jan 16;6:e21510. doi: 10.7554/eLife.21510. Elife. 2017. PMID: 28092261 Free PMC article.
-
Structure and function of spliceosomal DEAH-box ATPases.Biol Chem. 2023 Jul 17;404(8-9):851-866. doi: 10.1515/hsz-2023-0157. Print 2023 Jul 26. Biol Chem. 2023. PMID: 37441768 Review.
-
Structure, function and regulation of spliceosomal RNA helicases.Curr Opin Cell Biol. 2012 Jun;24(3):431-8. doi: 10.1016/j.ceb.2012.03.004. Epub 2012 Mar 29. Curr Opin Cell Biol. 2012. PMID: 22464735 Review.
Cited by
-
Continuous millisecond conformational cycle of a DEAH box helicase reveals control of domain motions by atomic-scale transitions.Commun Biol. 2023 Apr 7;6(1):379. doi: 10.1038/s42003-023-04751-z. Commun Biol. 2023. PMID: 37029280 Free PMC article.
-
Regulation of the DEAH/RHA helicase Prp43 by the G-patch factor Pfa1.Proc Natl Acad Sci U S A. 2022 Nov 29;119(48):e2203567119. doi: 10.1073/pnas.2203567119. Epub 2022 Nov 21. Proc Natl Acad Sci U S A. 2022. PMID: 36409901 Free PMC article.
-
Conformational dynamics of the RNA binding channel regulates loading and translocation of the DEAH-box helicase Prp43.Nucleic Acids Res. 2023 Jul 7;51(12):6430-6442. doi: 10.1093/nar/gkad362. Nucleic Acids Res. 2023. PMID: 37167006 Free PMC article.
-
Regulation of 3' splice site selection after step 1 of splicing by spliceosomal C* proteins.Sci Adv. 2023 Mar 3;9(9):eadf1785. doi: 10.1126/sciadv.adf1785. Epub 2023 Mar 3. Sci Adv. 2023. PMID: 36867703 Free PMC article.
-
Structural basis of catalytic activation in human splicing.Nature. 2023 May;617(7962):842-850. doi: 10.1038/s41586-023-06049-w. Epub 2023 May 10. Nature. 2023. PMID: 37165190 Free PMC article.
References
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
