

Perception of solar UV radiation by plants: photoreceptors and mechanisms
- PMID: 33826733
- PMCID: PMC8260113
- DOI: 10.1093/plphys/kiab162
Perception of solar UV radiation by plants: photoreceptors and mechanisms
Abstract
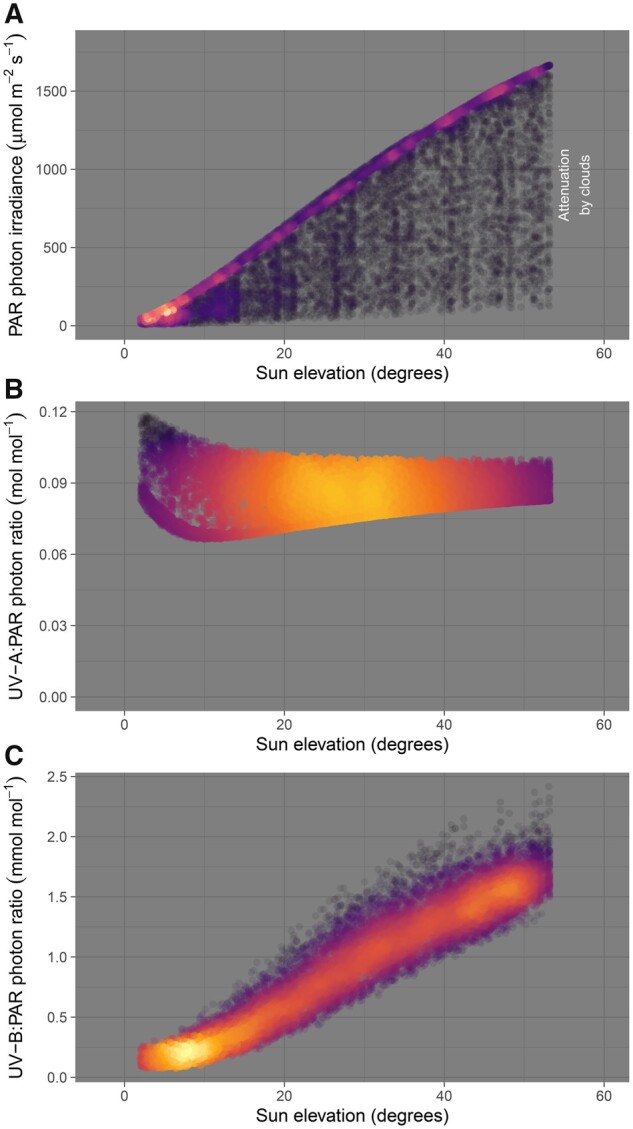
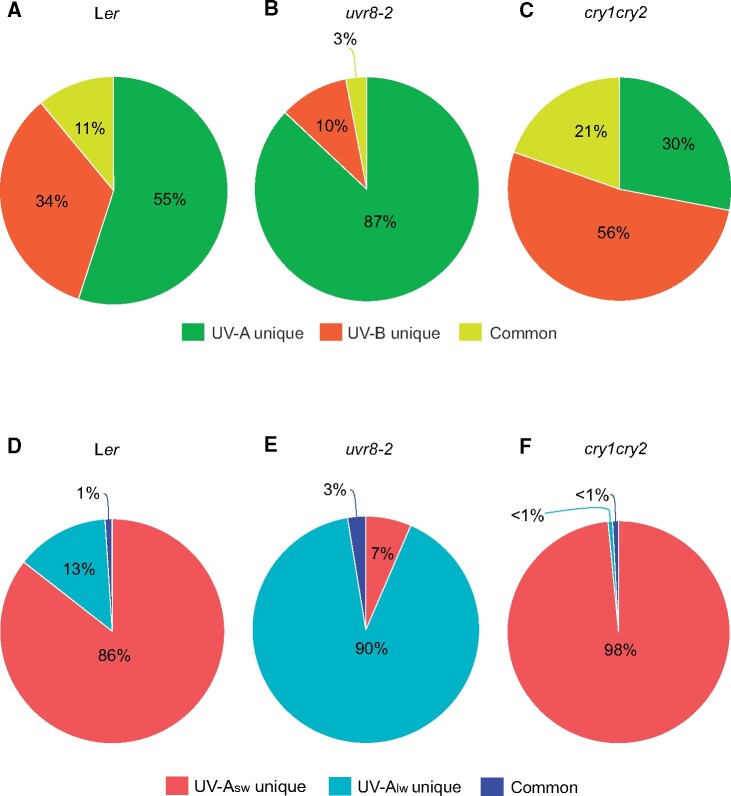
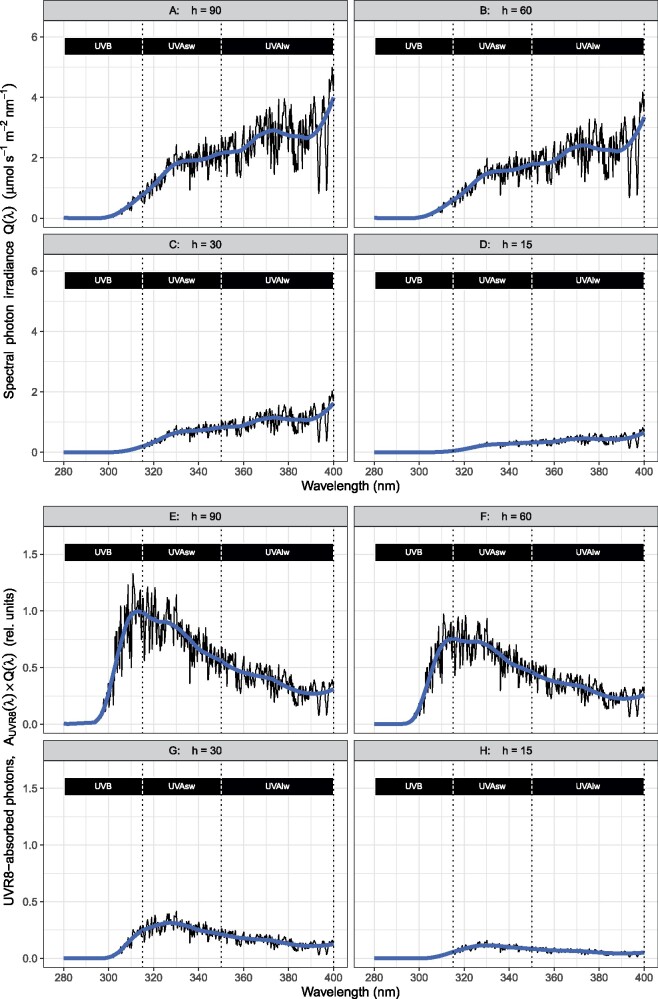
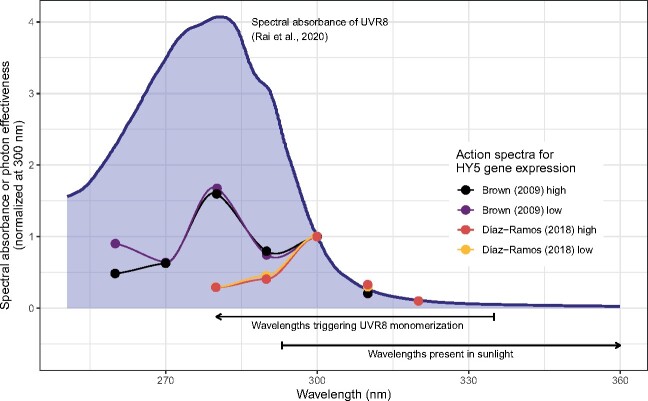
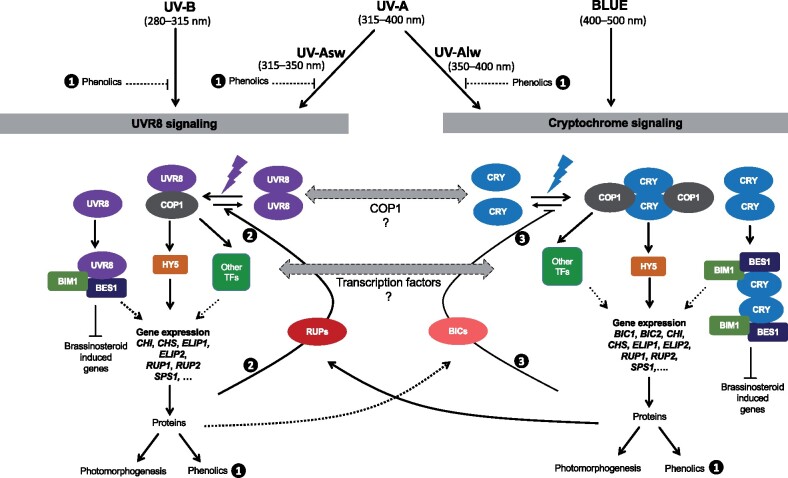
About 95% of the ultraviolet (UV) photons reaching the Earth's surface are UV-A (315-400 nm) photons. Plant responses to UV-A radiation have been less frequently studied than those to UV-B (280-315 nm) radiation. Most previous studies on UV-A radiation have used an unrealistic balance between UV-A, UV-B, and photosynthetically active radiation (PAR). Consequently, results from these studies are difficult to interpret from an ecological perspective, leaving an important gap in our understanding of the perception of solar UV radiation by plants. Previously, it was assumed UV-A/blue photoreceptors, cryptochromes and phototropins mediated photomorphogenic responses to UV-A radiation and "UV-B photoreceptor" UV RESISTANCE LOCUS 8 (UVR8) to UV-B radiation. However, our understanding of how UV-A radiation is perceived by plants has recently improved. Experiments using a realistic balance between UV-B, UV-A, and PAR have demonstrated that UVR8 can play a major role in the perception of both UV-B and short-wavelength UV-A (UV-Asw, 315 to ∼350 nm) radiation. These experiments also showed that UVR8 and cryptochromes jointly regulate gene expression through interactions that alter the relative sensitivity to UV-B, UV-A, and blue wavelengths. Negative feedback loops on the action of these photoreceptors can arise from gene expression, signaling crosstalk, and absorption of UV photons by phenolic metabolites. These interactions explain why exposure to blue light modulates photomorphogenic responses to UV-B and UV-Asw radiation. Future studies will need to distinguish between short and long wavelengths of UV-A radiation and to consider UVR8's role as a UV-B/UV-Asw photoreceptor in sunlight.
© The Author(s) 2021. Published by Oxford University Press on behalf of American Society of Plant Biologists.
Figures
References
-
- Aphalo PJ, Albert A, McLeod AR, Heikkilä A, Gómez I, López Figueroa F, Robson TM, Strid Å (2012) Manipulating UV radiation. InAphalo PJ, Albert A, Björn LO, McLeod AR, Robson TM, Rosenqvist E, eds, Beyond the Visible: A Handbook of Best Practice in Plant UV Photobiology , Ed 1,University of Helsinki, Department of Biosciences, Division of Plant Biology, pp 35–70
-
- Aphalo PJ (2015) The r4photobiology suite: spectral irradiance. UV4Plants Bull 1:21–29
-
- Ballaré CL (2014) Light regulation of plant defense. Annu Rev Plant Biol 65:335–363 - PubMed
-
- Ballaré CL, Pierik R (2017) The shade-avoidance syndrome: Multiple signals and ecological consequences. Plant Cell Environ 40:2530–2543 - PubMed
-
- Banerjee R, Schleicher E, Meier S, Viana RM, Pokorny R, Ahmad M, Bittl R, Batschauer A (2007) The signaling state of Arabidopsis cryptochrome 2 contains flavin semiquinone. J Biol Chem 282:14916–14922 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
