

Clustering and Erratic Movement Patterns of Syringe-Injected versus Mosquito-Inoculated Malaria Sporozoites Underlie Decreased Infectivity
- PMID: 33827910
- PMCID: PMC8546700
- DOI: 10.1128/mSphere.00218-21
Clustering and Erratic Movement Patterns of Syringe-Injected versus Mosquito-Inoculated Malaria Sporozoites Underlie Decreased Infectivity
Abstract
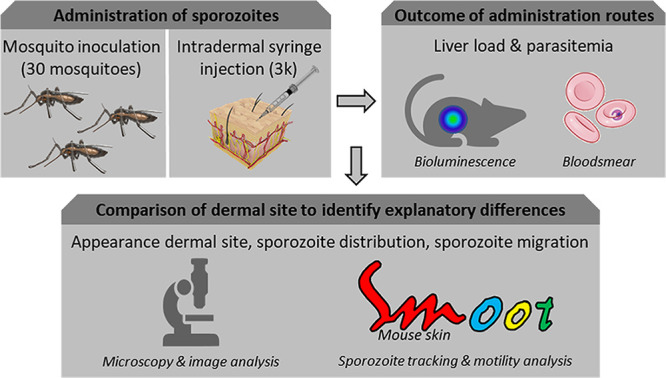
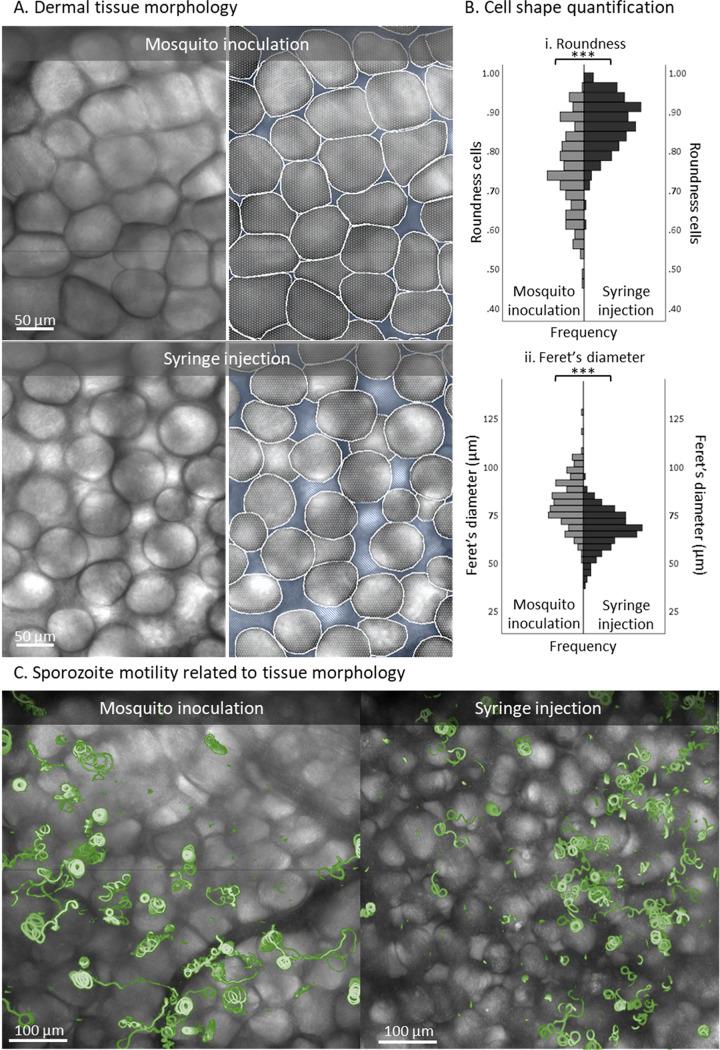
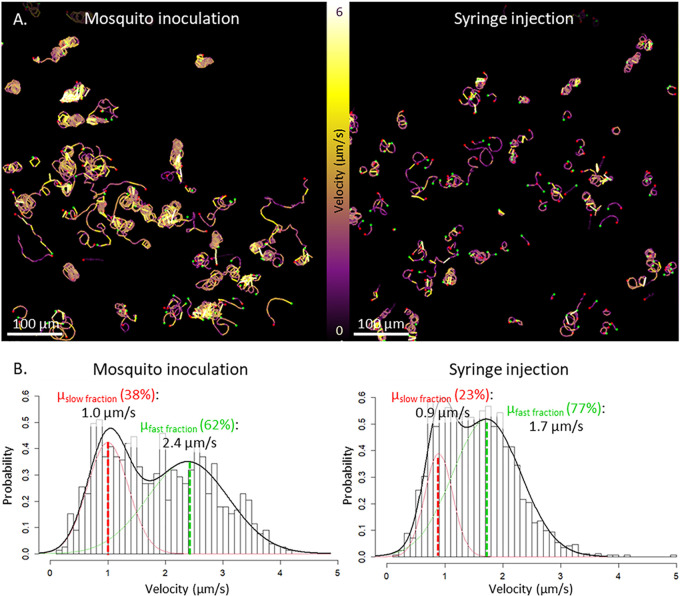
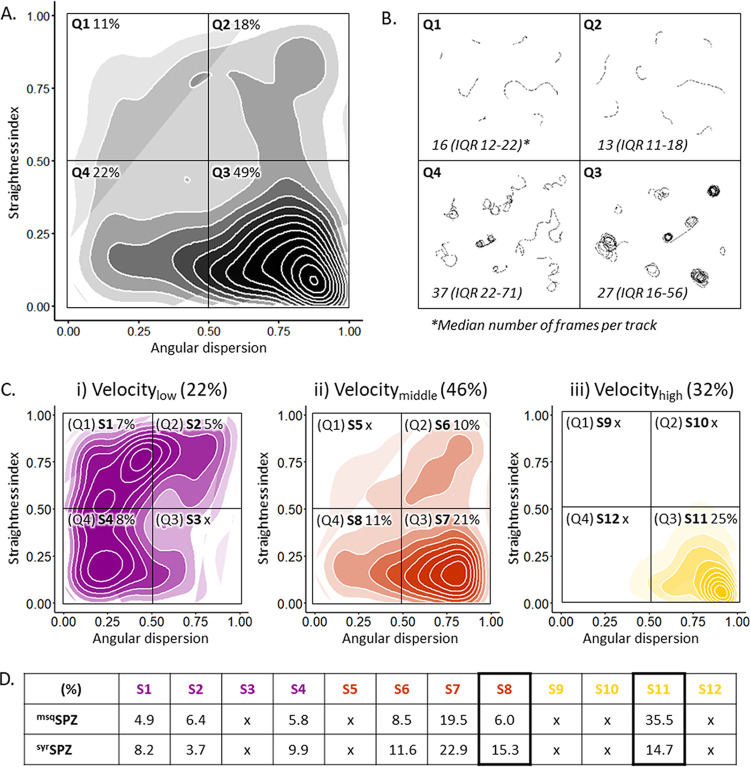
Malaria vaccine candidates based on live, attenuated sporozoites have led to high levels of protection. However, their efficacy critically depends on the sporozoites' ability to reach and infect the host liver. Administration via mosquito inoculation is by far the most potent method for inducing immunity but highly impractical. Here, we observed that intradermal syringe-injected Plasmodium berghei sporozoites (syrSPZ) were 3-fold less efficient in migrating to and infecting mouse liver than mosquito-inoculated sporozoites (msqSPZ). This was related to a clustered dermal distribution (2-fold-decreased median distance between syrSPZ and msqSPZ) and, more importantly, a 1.4-fold (significantly)-slower and more erratic movement pattern. These erratic movement patterns were likely caused by alteration of dermal tissue morphology (>15-μm intercellular gaps) due to injection of fluid and may critically decrease sporozoite infectivity. These results suggest that novel microvolume-based administration technologies hold promise for replicating the success of mosquito-inoculated live, attenuated sporozoite vaccines.IMPORTANCE Malaria still causes a major burden on global health and the economy. The efficacy of live, attenuated malaria sporozoites as vaccine candidates critically depends on their ability to migrate to and infect the host liver. This work sheds light on the effect of different administration routes on sporozoite migration. We show that the delivery of sporozoites via mosquito inoculation is more efficient than syringe injection; however, this route of administration is highly impractical for vaccine purposes. Using confocal microscopy and automated imaging software, we demonstrate that syringe-injected sporozoites do cluster, move more slowly, and display more erratic movement due to alterations in tissue morphology. These findings indicate that microneedle-based engineering solutions hold promise for replicating the success of mosquito-inoculated live, attenuated sporozoite vaccines.
Keywords: Plasmodium berghei; fluorescent microscopy; malaria; motility; sporozoite.
Copyright © 2021 de Korne et al.
Figures



Similar articles
-
Infectivity of Plasmodium berghei sporozoites delivered by intravenous inoculation versus mosquito bite: implications for sporozoite vaccine trials.Infect Immun. 1999 Aug;67(8):4285-9. doi: 10.1128/IAI.67.8.4285-4289.1999. Infect Immun. 1999. PMID: 10417207 Free PMC article.
-
Intradermal immunization of mice with radiation-attenuated sporozoites of Plasmodium yoelii induces effective protective immunity.Malar J. 2010 Dec 15;9:362. doi: 10.1186/1475-2875-9-362. Malar J. 2010. PMID: 21159170 Free PMC article.
-
Comparison of Plasmodium berghei challenge models for the evaluation of pre-erythrocytic malaria vaccines and their effect on perceived vaccine efficacy.Malar J. 2010 May 27;9:145. doi: 10.1186/1475-2875-9-145. Malar J. 2010. PMID: 20507620 Free PMC article.
-
Route of administration of attenuated sporozoites is instrumental in rendering immunity against Plasmodia infection.Vaccine. 2016 Jun 14;34(28):3229-34. doi: 10.1016/j.vaccine.2016.04.095. Epub 2016 May 6. Vaccine. 2016. PMID: 27160038 Review.
-
The whole parasite, pre-erythrocytic stage approach to malaria vaccine development: a review.Curr Opin Infect Dis. 2013 Oct;26(5):420-8. doi: 10.1097/QCO.0000000000000002. Curr Opin Infect Dis. 2013. PMID: 23982233 Review.
Cited by
-
Malaria: influence of Anopheles mosquito saliva on Plasmodium infection.Trends Immunol. 2023 Apr;44(4):256-265. doi: 10.1016/j.it.2023.02.005. Epub 2023 Mar 22. Trends Immunol. 2023. PMID: 36964020 Free PMC article. Review.
-
Sporozoite motility as a quantitative readout for anti-CSP antibody inhibition.Sci Rep. 2022 Oct 13;12(1):17194. doi: 10.1038/s41598-022-22154-8. Sci Rep. 2022. PMID: 36229488 Free PMC article.
-
Ultra-low volume intradermal administration of radiation-attenuated sporozoites with the glycolipid adjuvant 7DW8-5 completely protects mice against malaria.Sci Rep. 2024 Feb 4;14(1):2881. doi: 10.1038/s41598-024-53118-9. Sci Rep. 2024. PMID: 38311678 Free PMC article.
-
Plasmodium sporozoite shows distinct motility patterns in responses to three-dimensional environments.iScience. 2024 Jul 5;27(8):110463. doi: 10.1016/j.isci.2024.110463. eCollection 2024 Aug 16. iScience. 2024. PMID: 39129829 Free PMC article.
References
-
- Richie TL, Billingsley PF, Sim BK, James ER, Chakravarty S, Epstein JE, Lyke KE, Mordmuller B, Alonso P, Duffy PE, Doumbo OK, Sauerwein RW, Tanner M, Abdulla S, Kremsner PG, Seder RA, Hoffman SL. 2015. Progress with Plasmodium falciparum sporozoite (PfSPZ)-based malaria vaccines. Vaccine 33:7452–7461. doi:10.1016/j.vaccine.2015.09.096. - DOI - PMC - PubMed
-
- Hoffman SL, Goh LM, Luke TC, Schneider I, Le TP, Doolan DL, Sacci J, de la Vega P, Dowler M, Paul C, Gordon DM, Stoute JA, Church LW, Sedegah M, Heppner DG, Ballou WR, Richie TL. 2002. Protection of humans against malaria by immunization with radiation-attenuated Plasmodium falciparum sporozoites. J Infect Dis 185:1155–1164. doi:10.1086/339409. - DOI - PubMed
-
- Epstein JE, Tewari K, Lyke KE, Sim BK, Billingsley PF, Laurens MB, Gunasekera A, Chakravarty S, James ER, Sedegah M, Richman A, Velmurugan S, Reyes S, Li M, Tucker K, Ahumada A, Ruben AJ, Li T, Stafford R, Eappen AG, Tamminga C, Bennett JW, Ockenhouse CF, Murphy JR, Komisar J, Thomas N, Loyevsky M, Birkett A, Plowe CV, Loucq C, Edelman R, Richie TL, Seder RA, Hoffman SL. 2011. Live attenuated malaria vaccine designed to protect through hepatic CD8(+) T cell immunity. Science 334:475–480. doi:10.1126/science.1211548. - DOI - PubMed
-
- Seder RA, Chang LJ, Enama ME, Zephir KL, Sarwar UN, Gordon IJ, Holman LA, James ER, Billingsley PF, Gunasekera A, Richman A, Chakravarty S, Manoj A, Velmurugan S, Li M, Ruben AJ, Li T, Eappen AG, Stafford RE, Plummer SH, Hendel CS, Novik L, Costner PJ, Mendoza FH, Saunders JG, Nason MC, Richardson JH, Murphy J, Davidson SA, Richie TL, Sedegah M, Sutamihardja A, Fahle GA, Lyke KE, Laurens MB, Roederer M, Tewari K, Epstein JE, Sim BK, Ledgerwood JE, Graham BS, Hoffman SL, Team VRCS, VRC 312 Study Team. 2013. Protection against malaria by intravenous immunization with a nonreplicating sporozoite vaccine. Science 341:1359–1365. doi:10.1126/science.1241800. - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
