

Type III-A CRISPR immunity promotes mutagenesis of staphylococci
- PMID: 33828299
- PMCID: PMC8820005
- DOI: 10.1038/s41586-021-03440-3
Type III-A CRISPR immunity promotes mutagenesis of staphylococci
Abstract
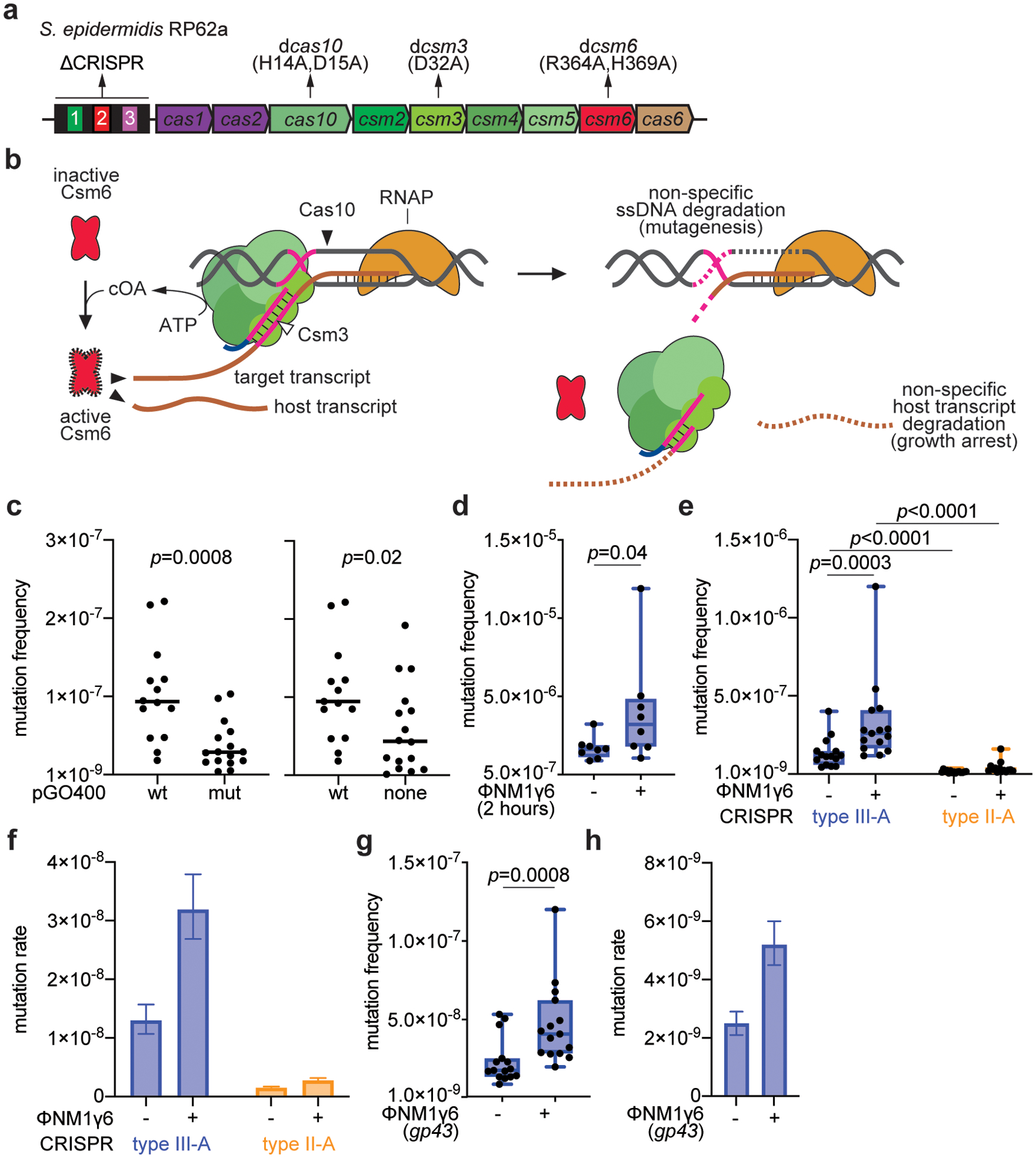
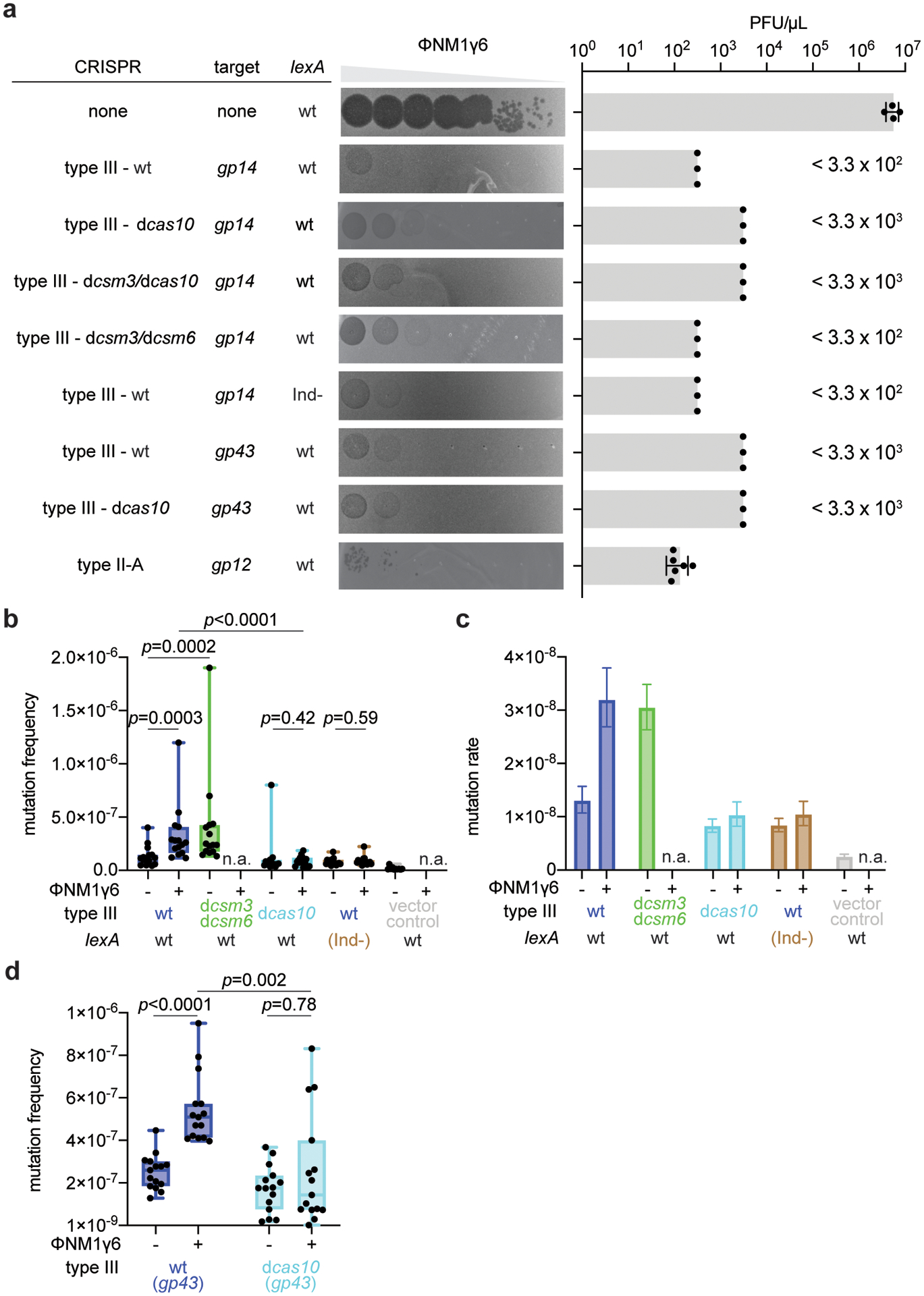
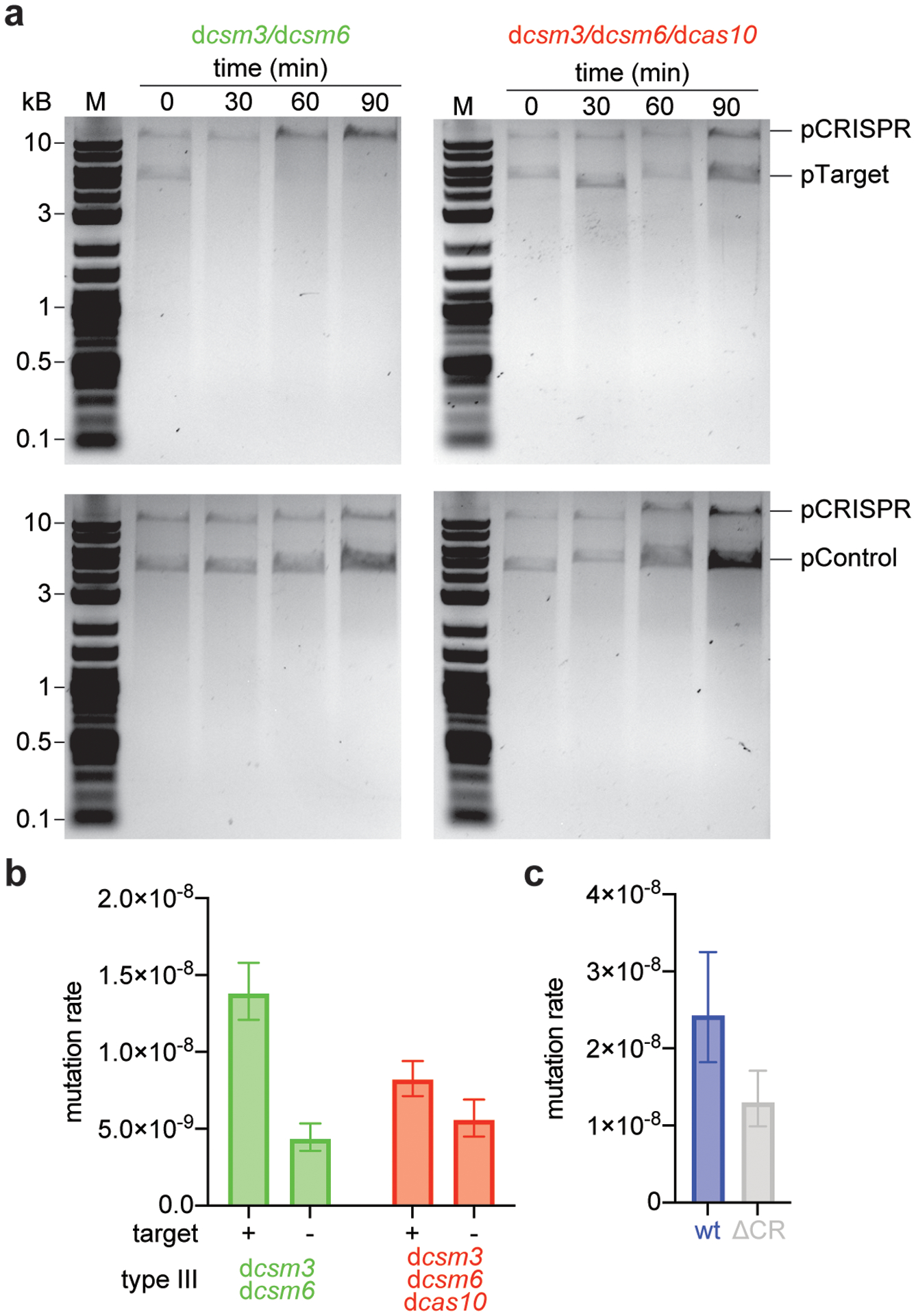
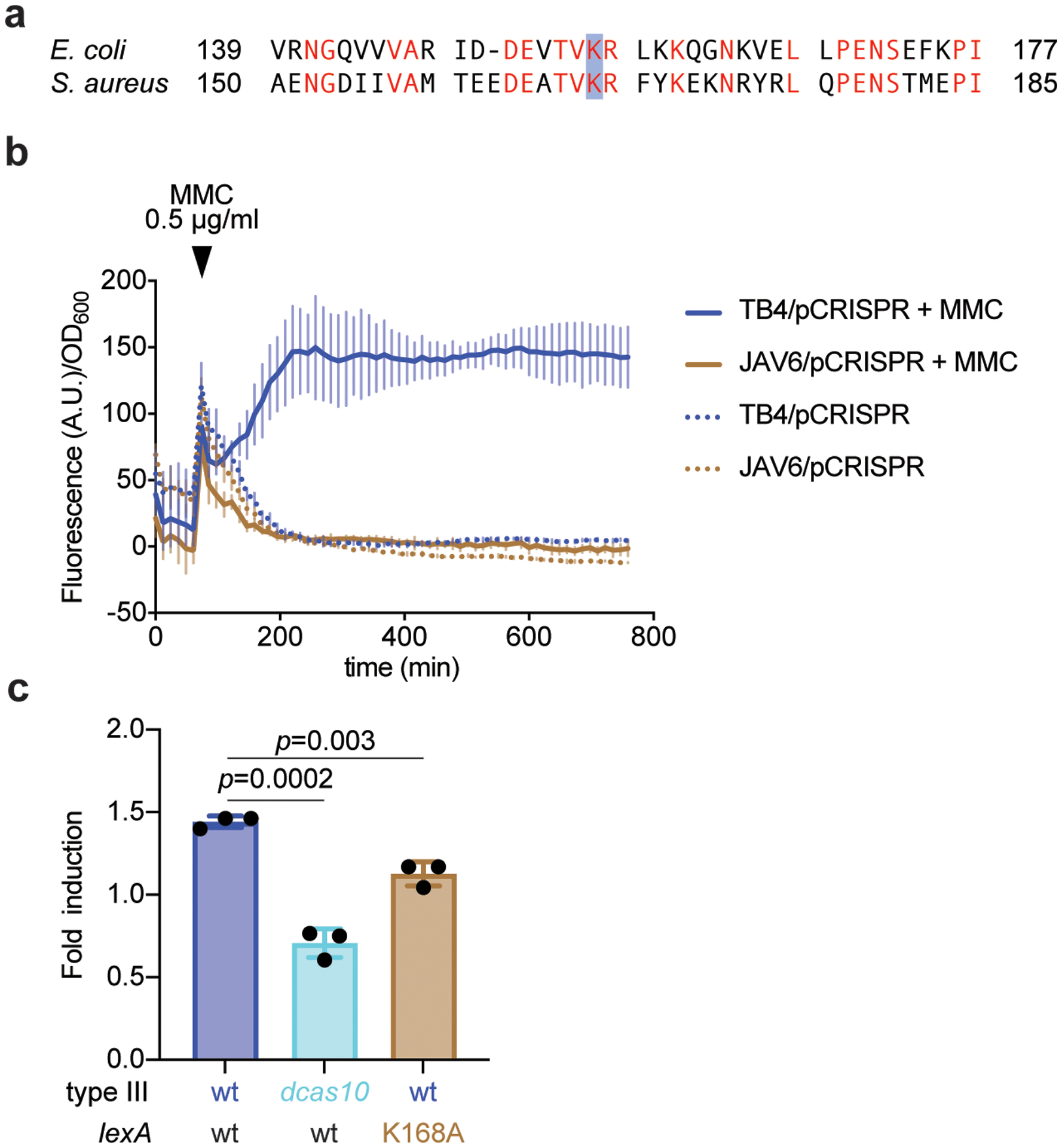
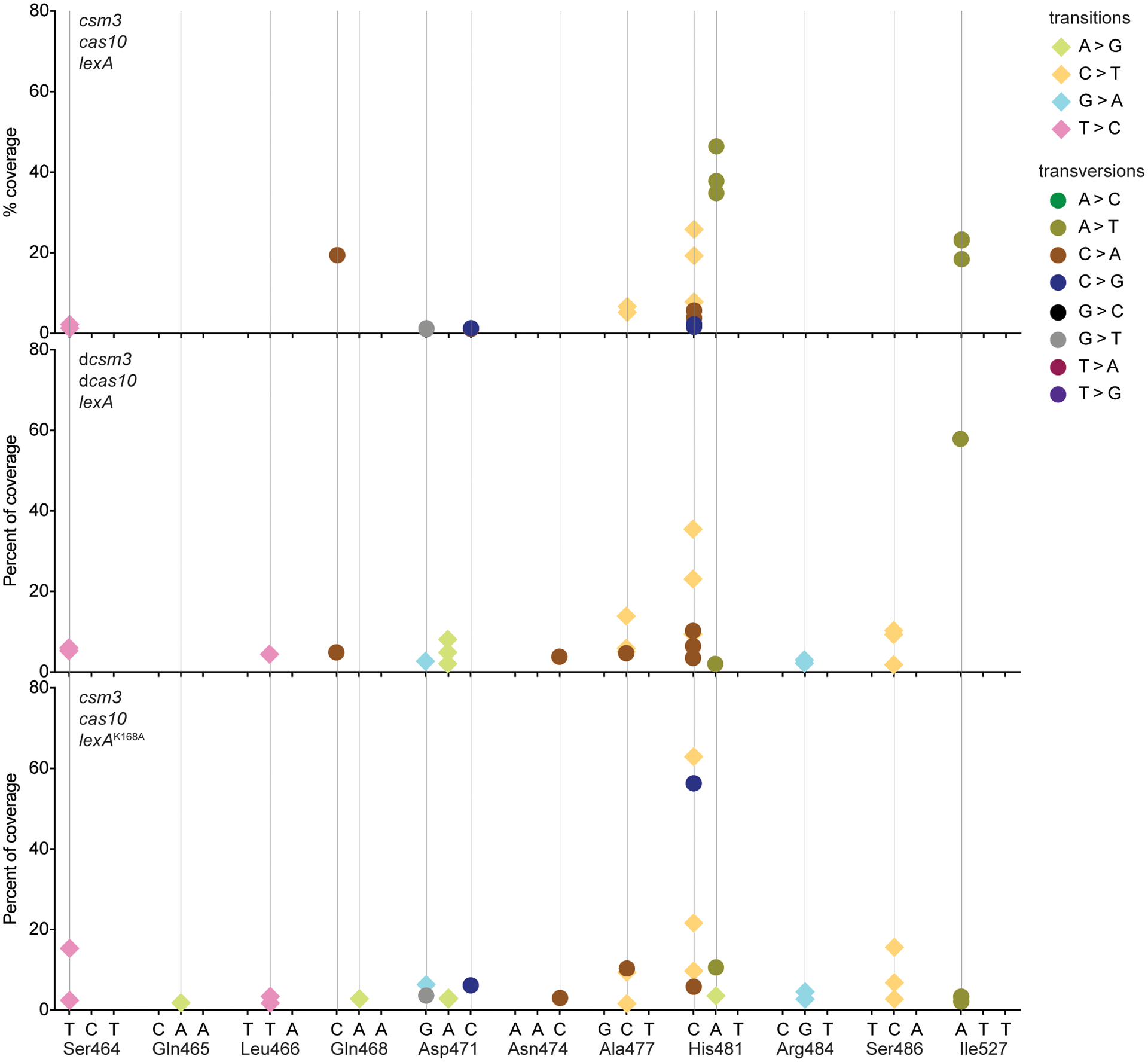
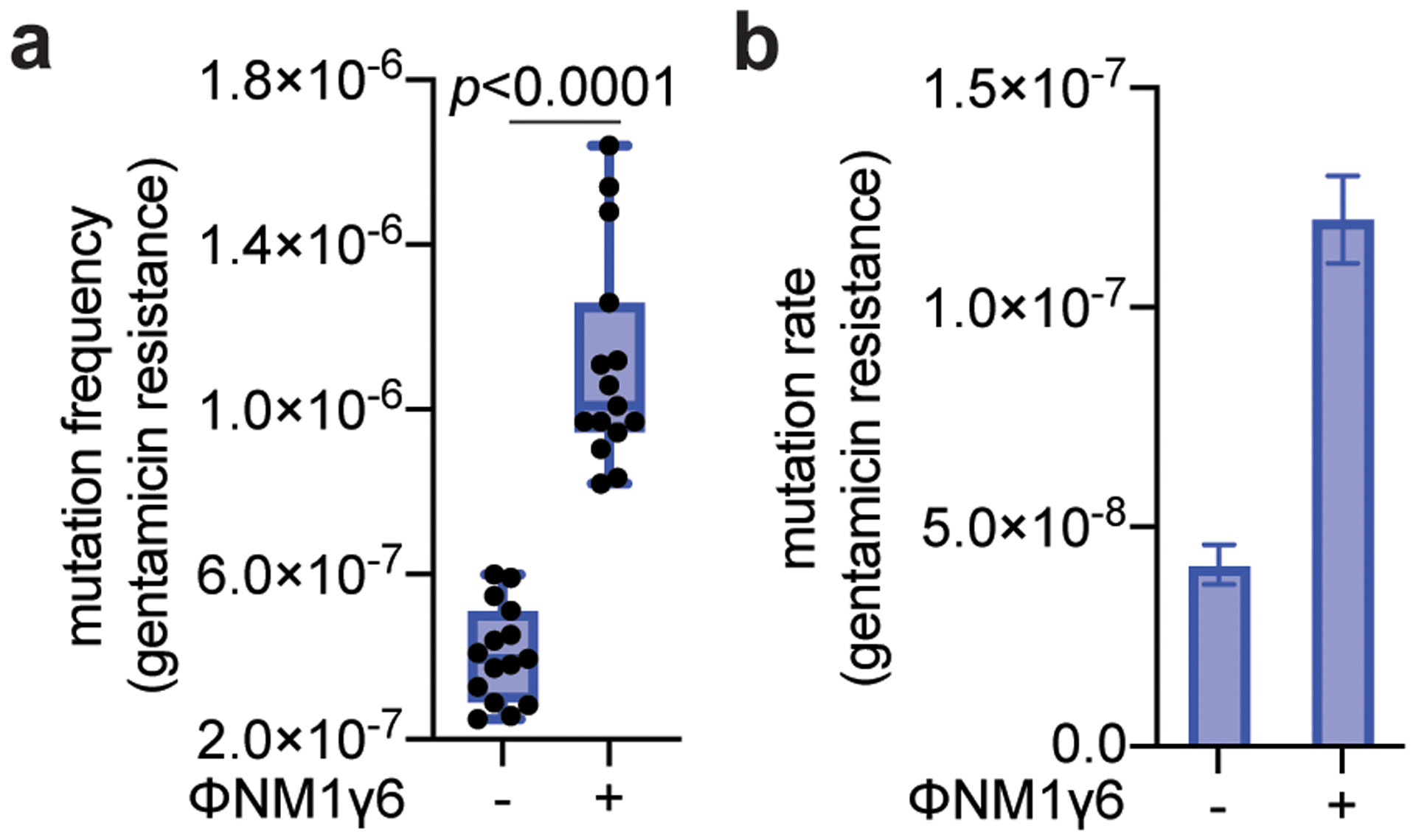
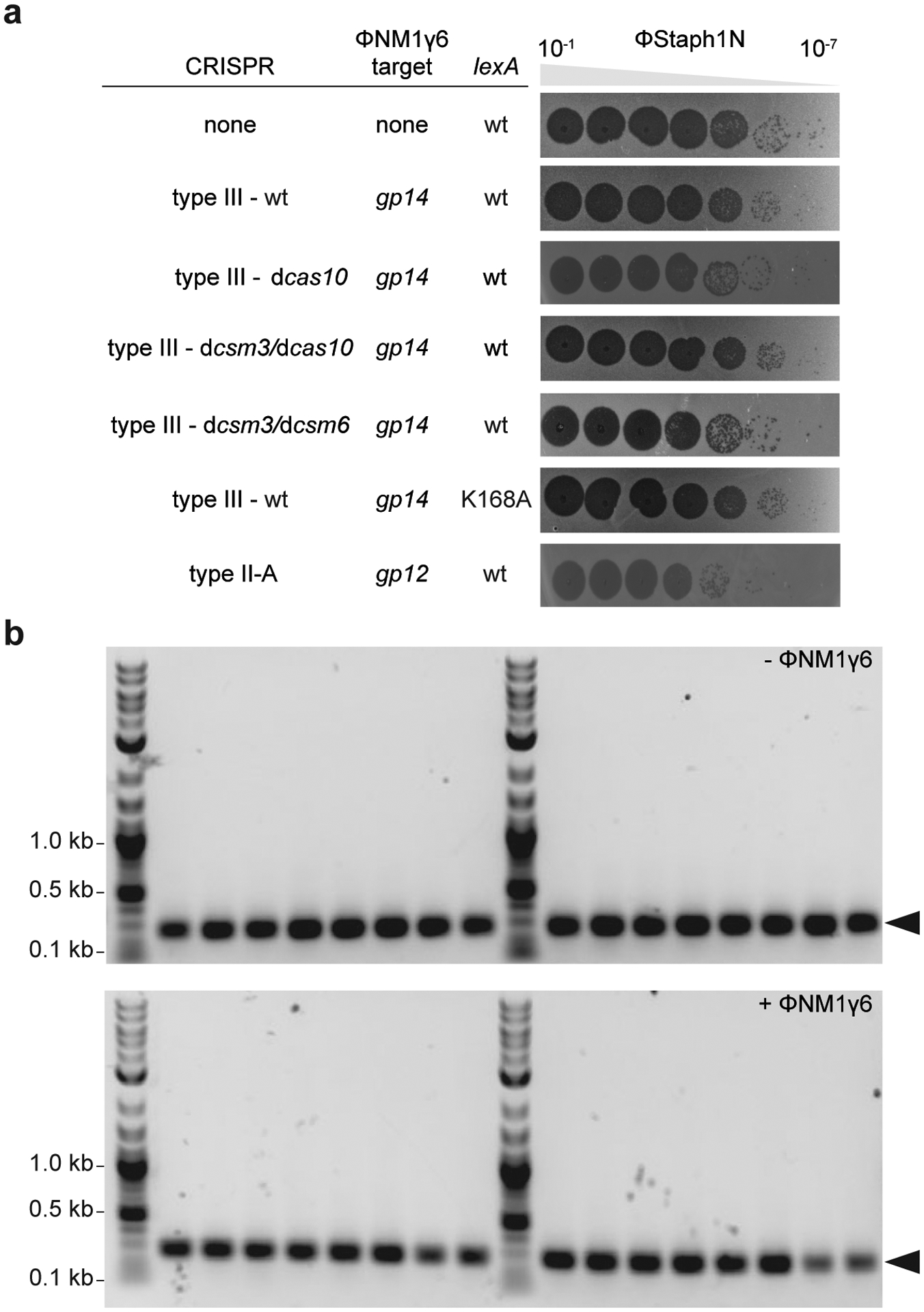
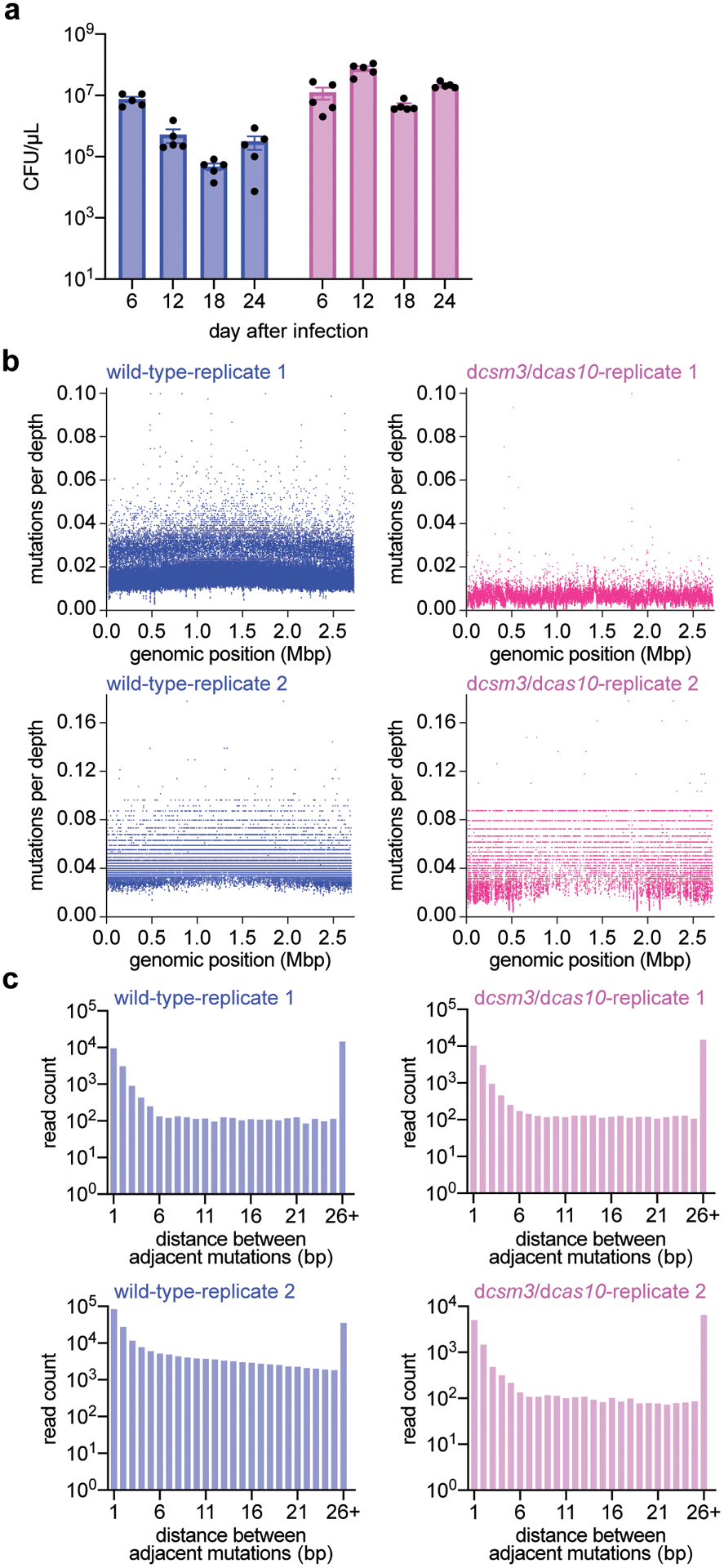
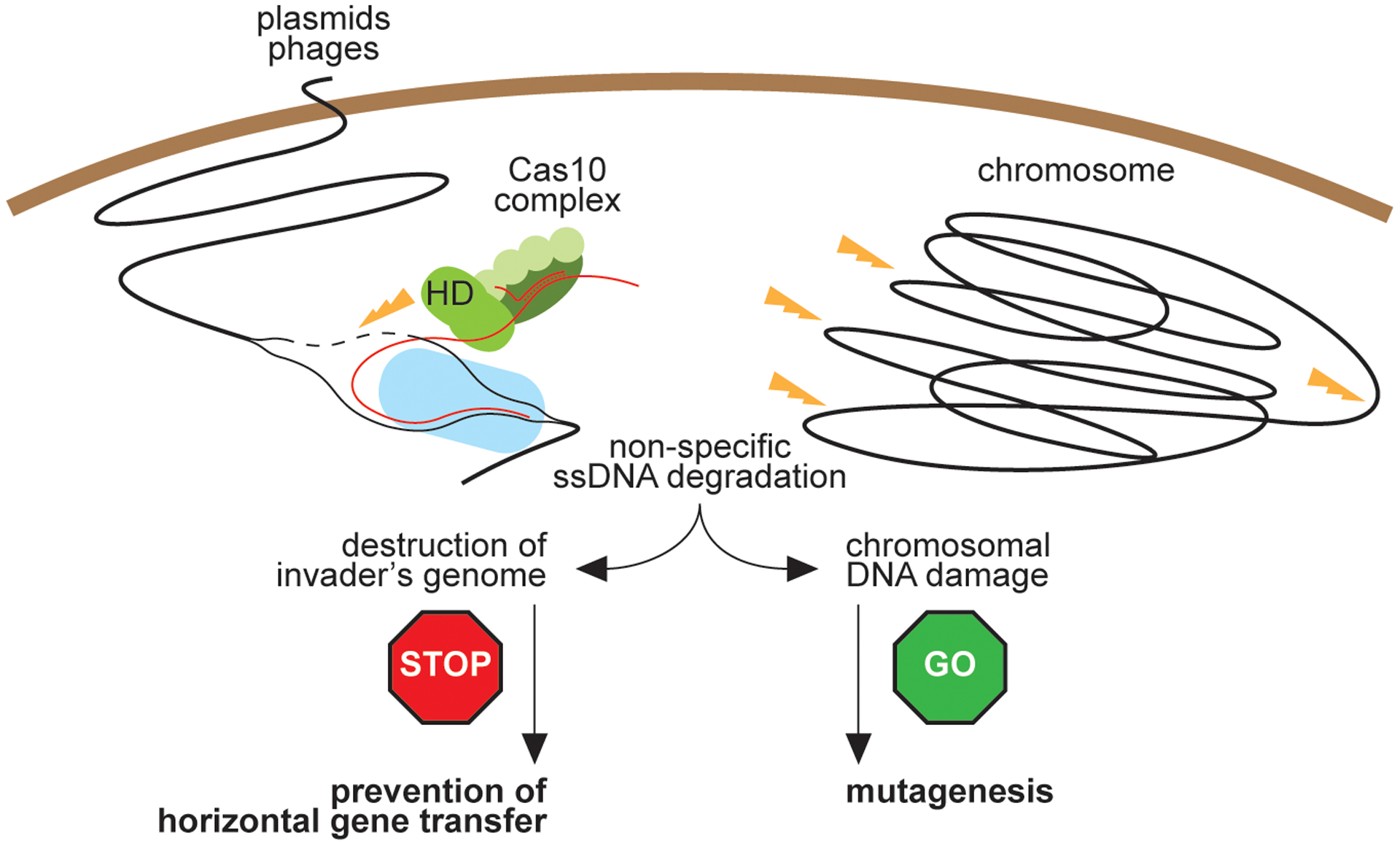
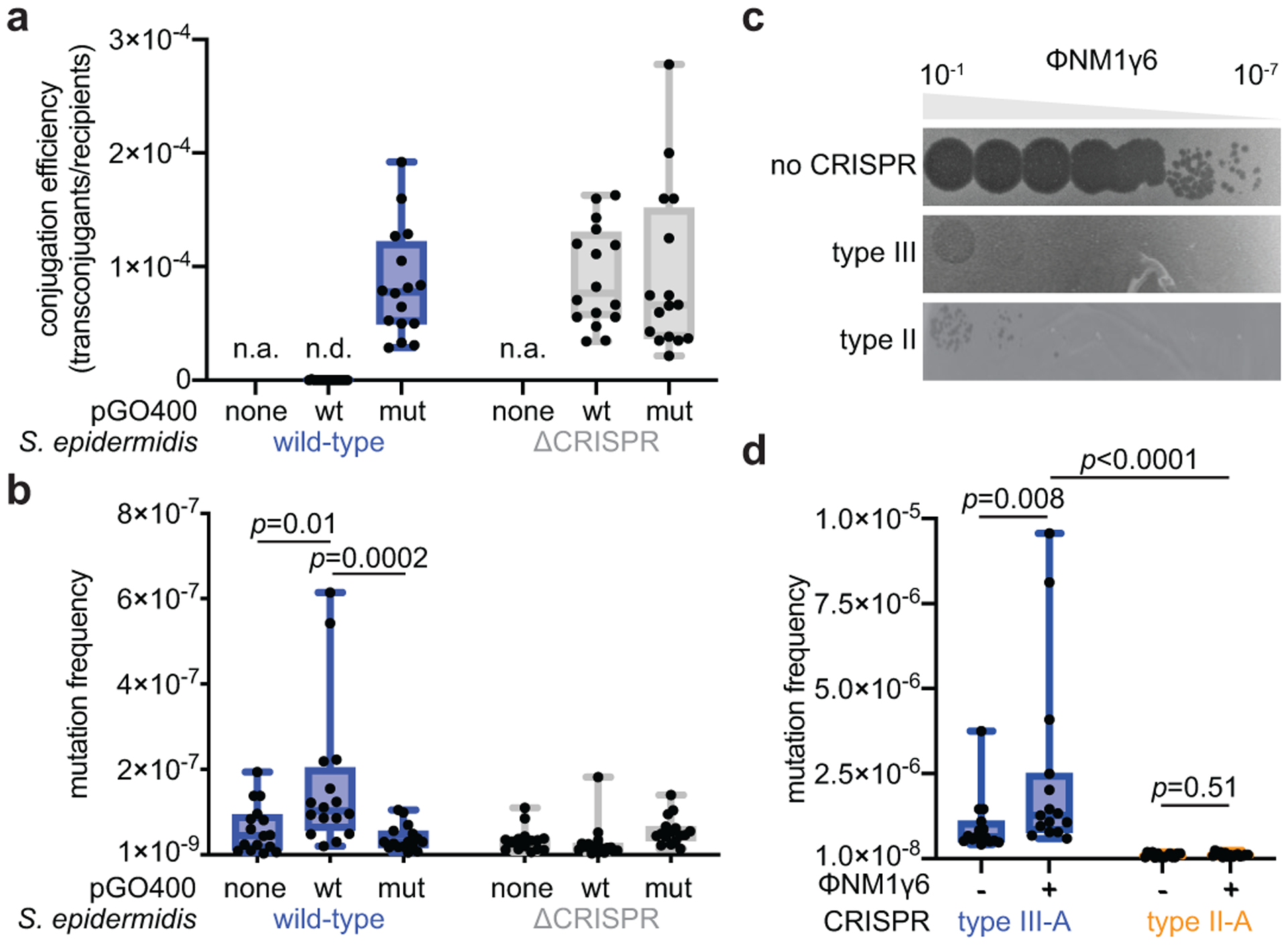
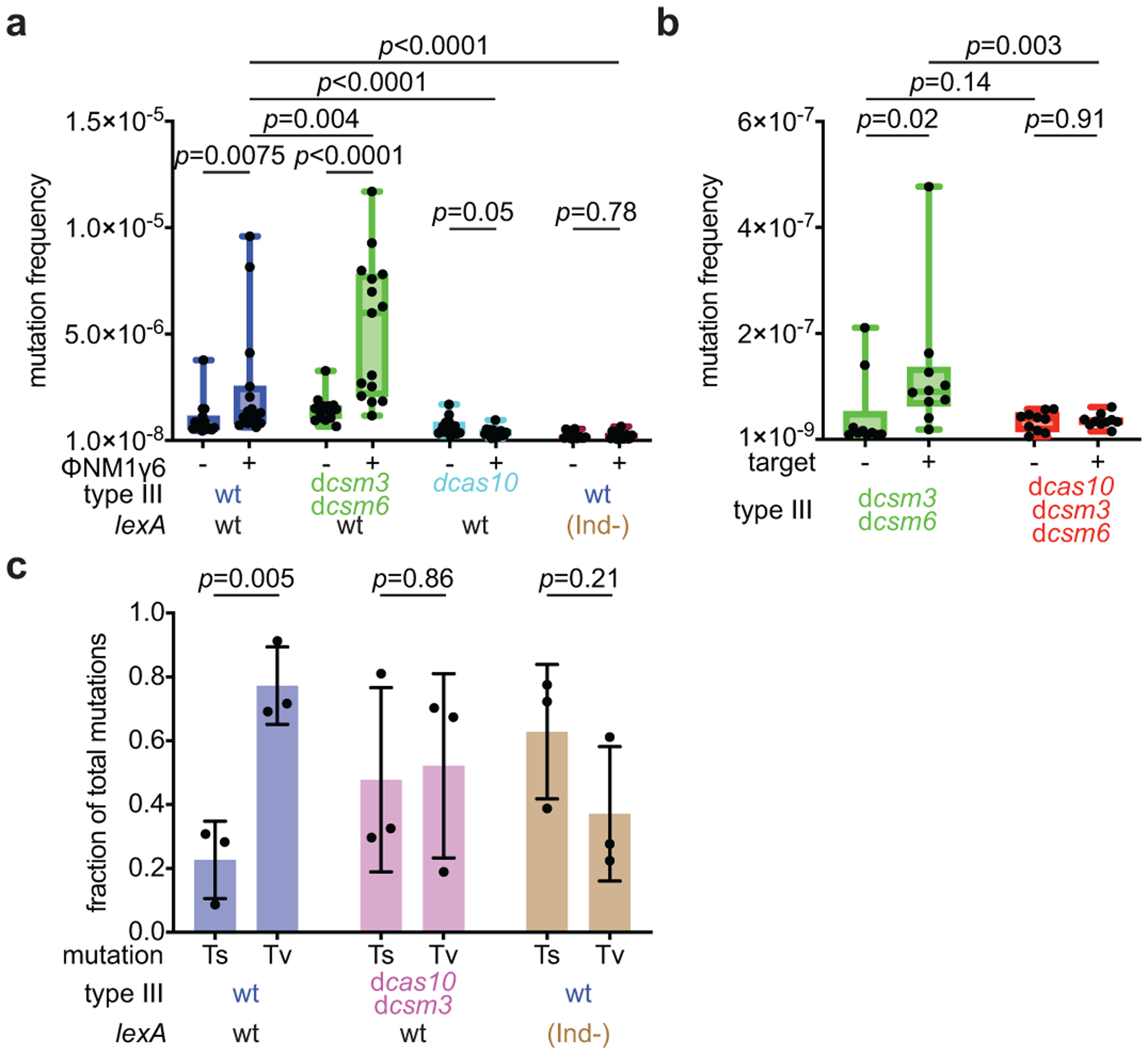
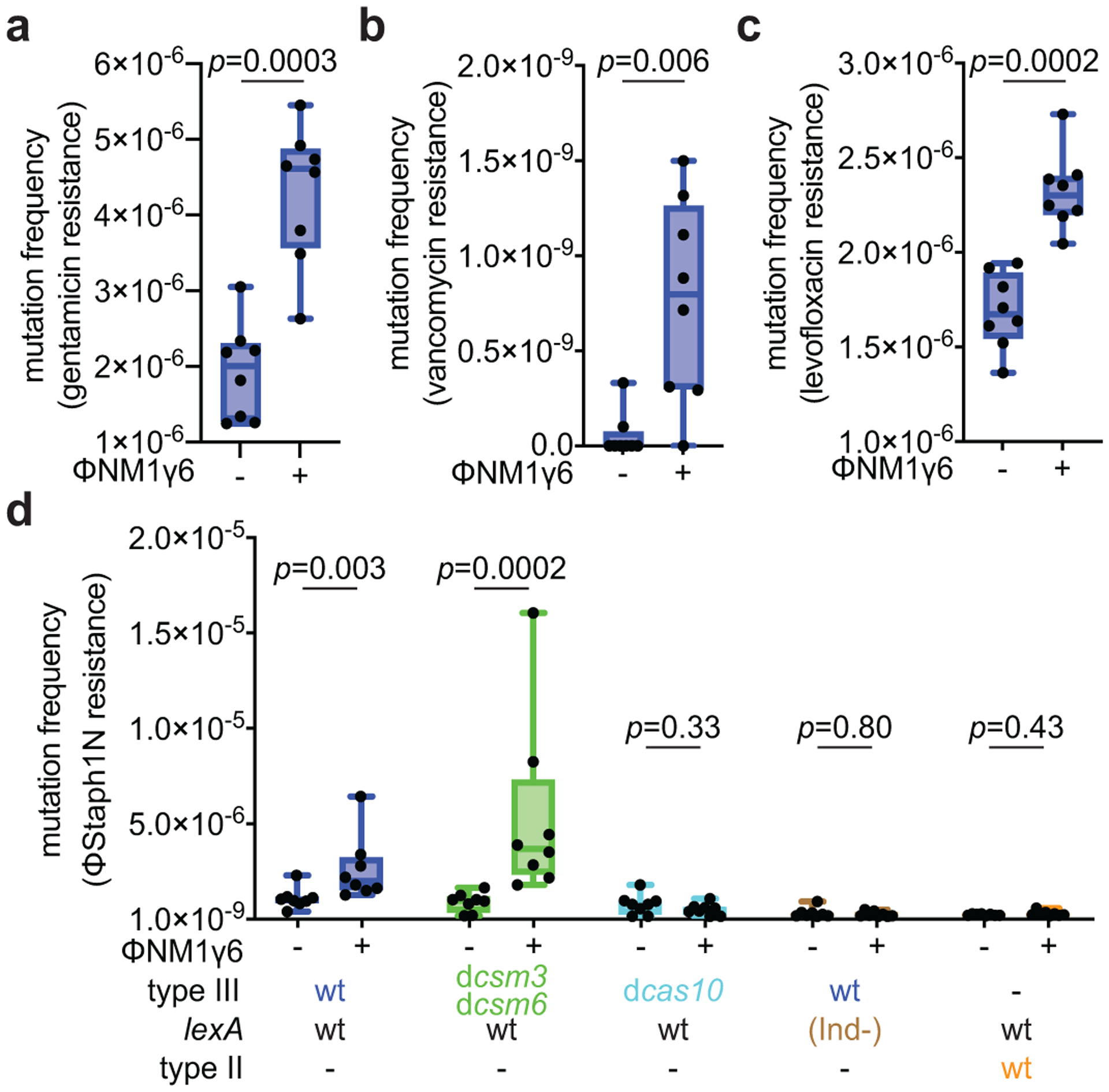
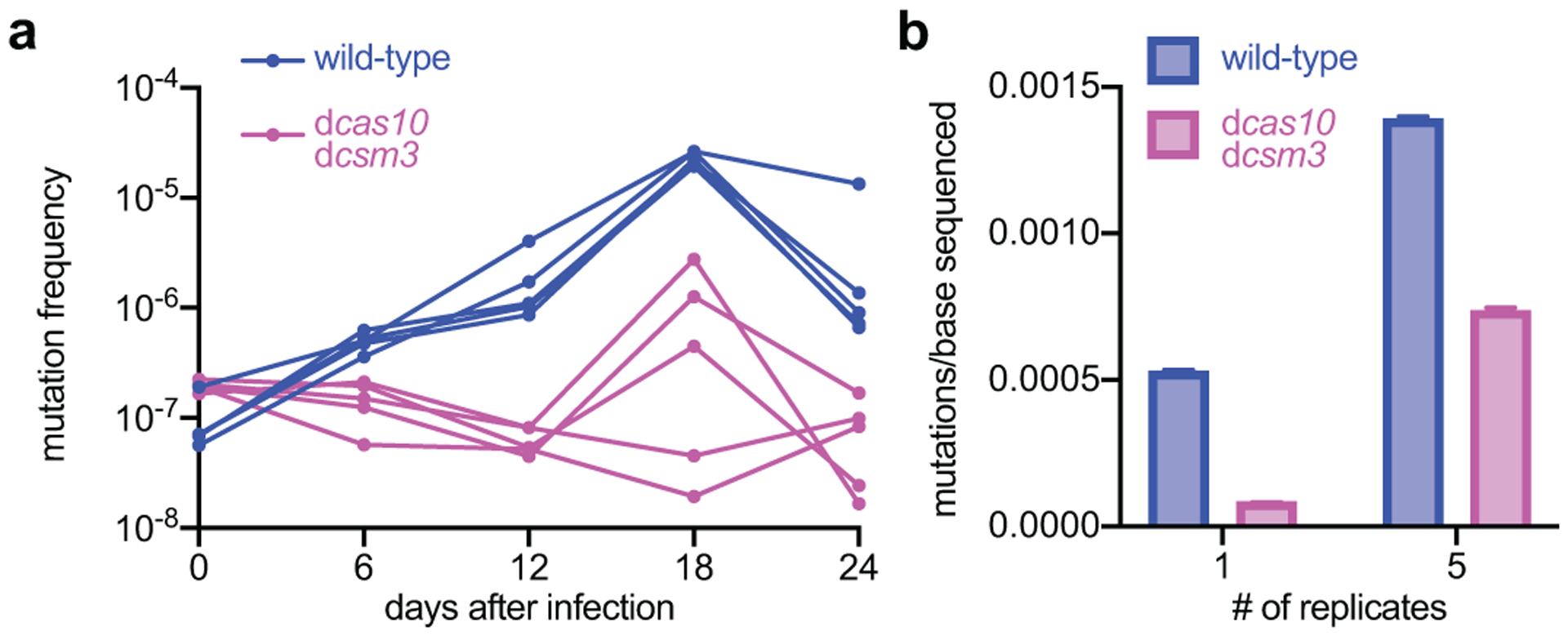
Horizontal gene transfer and mutation are the two major drivers of microbial evolution that enable bacteria to adapt to fluctuating environmental stressors1. Clustered, regularly interspaced, short palindromic repeats (CRISPR) systems use RNA-guided nucleases to direct sequence-specific destruction of the genomes of mobile genetic elements that mediate horizontal gene transfer, such as conjugative plasmids2 and bacteriophages3, thus limiting the extent to which bacteria can evolve by this mechanism. A subset of CRISPR systems also exhibit non-specific degradation of DNA4,5; however, whether and how this feature affects the host has not yet been examined. Here we show that the non-specific DNase activity of the staphylococcal type III-A CRISPR-Cas system increases mutations in the host and accelerates the generation of antibiotic resistance in Staphylococcus aureus and Staphylococcus epidermidis. These mutations require the induction of the SOS response to DNA damage and display a distinct pattern. Our results demonstrate that by differentially affecting both mechanisms that generate genetic diversity, type III-A CRISPR systems can modulate the evolution of the bacterial host.
Conflict of interest statement
Figures
References
-
- Barrangou R et al. CRISPR provides acquired resistance against viruses in prokaryotes. Science 315, 1709–1712. - PubMed
-
- Kazlauskiene M, Tamulaitis G, Kostiuk G, Venclovas C & Siksnys V Spatiotemporal Control of Type III-A CRISPR-Cas Immunity: Coupling DNA Degradation with the Target RNA Recognition. Mol. Cell 62, 295–306. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
