

A human coronavirus evolves antigenically to escape antibody immunity
- PMID: 33831132
- PMCID: PMC8031418
- DOI: 10.1371/journal.ppat.1009453
A human coronavirus evolves antigenically to escape antibody immunity
Abstract
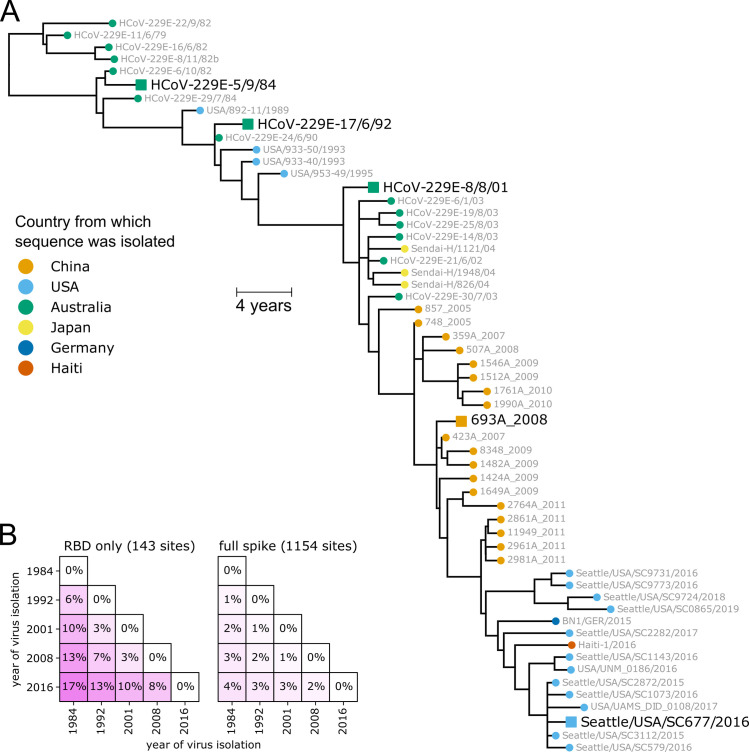
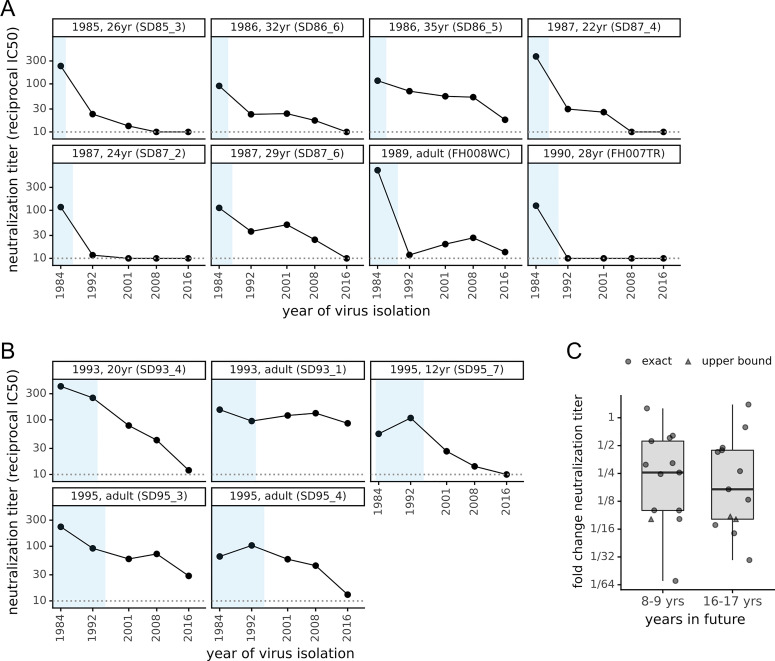
There is intense interest in antibody immunity to coronaviruses. However, it is unknown if coronaviruses evolve to escape such immunity, and if so, how rapidly. Here we address this question by characterizing the historical evolution of human coronavirus 229E. We identify human sera from the 1980s and 1990s that have neutralizing titers against contemporaneous 229E that are comparable to the anti-SARS-CoV-2 titers induced by SARS-CoV-2 infection or vaccination. We test these sera against 229E strains isolated after sera collection, and find that neutralizing titers are lower against these "future" viruses. In some cases, sera that neutralize contemporaneous 229E viral strains with titers >1:100 do not detectably neutralize strains isolated 8-17 years later. The decreased neutralization of "future" viruses is due to antigenic evolution of the viral spike, especially in the receptor-binding domain. If these results extrapolate to other coronaviruses, then it may be advisable to periodically update SARS-CoV-2 vaccines.
Conflict of interest statement
I have read the journal’s policy and the authors of this manuscript have the following competing interests: MJB has consulted for Moderna and Vir Biotechnologies, and received research funding from Regeneron and Vir Biotechnologies. JAE has consulted for Meissa Vaccines and Sanofi Pasteur, and received research funding from Merck, GlaxoSmithKline, Pfizer, and AstraZeneca. The other authors declare no competing interests.
Figures
Comment in
-
Blast from the past for coronavirus immunity.Nat Rev Microbiol. 2021 Jun;19(6):344. doi: 10.1038/s41579-021-00566-z. Nat Rev Microbiol. 2021. PMID: 33883721 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
