

Phenotypic Differences between the Alzheimer's Disease-Related hAPP-J20 Model and Heterozygous Zbtb20 Knock-Out Mice
- PMID: 33833046
- PMCID: PMC8121260
- DOI: 10.1523/ENEURO.0089-21.2021
Phenotypic Differences between the Alzheimer's Disease-Related hAPP-J20 Model and Heterozygous Zbtb20 Knock-Out Mice
Abstract
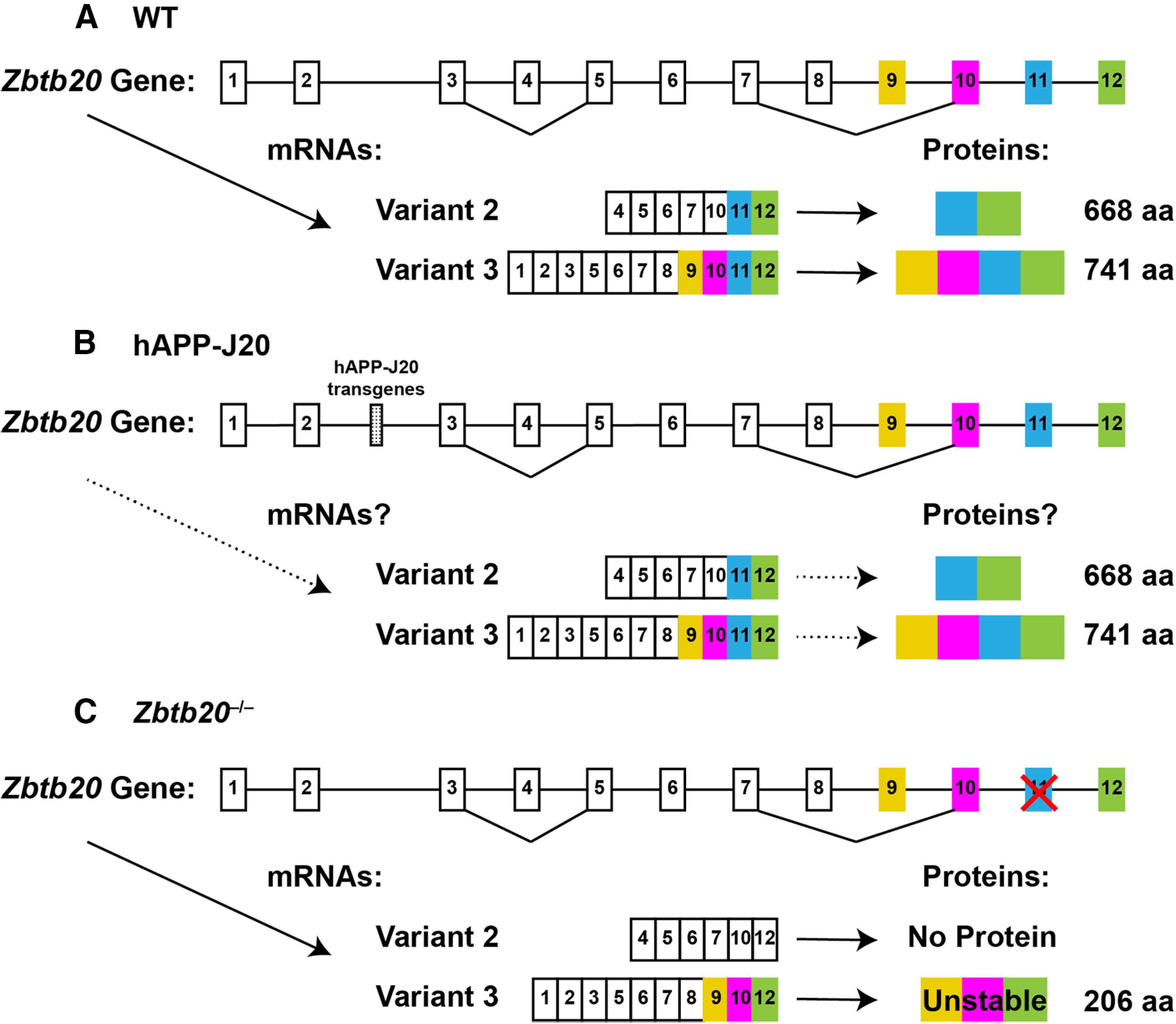
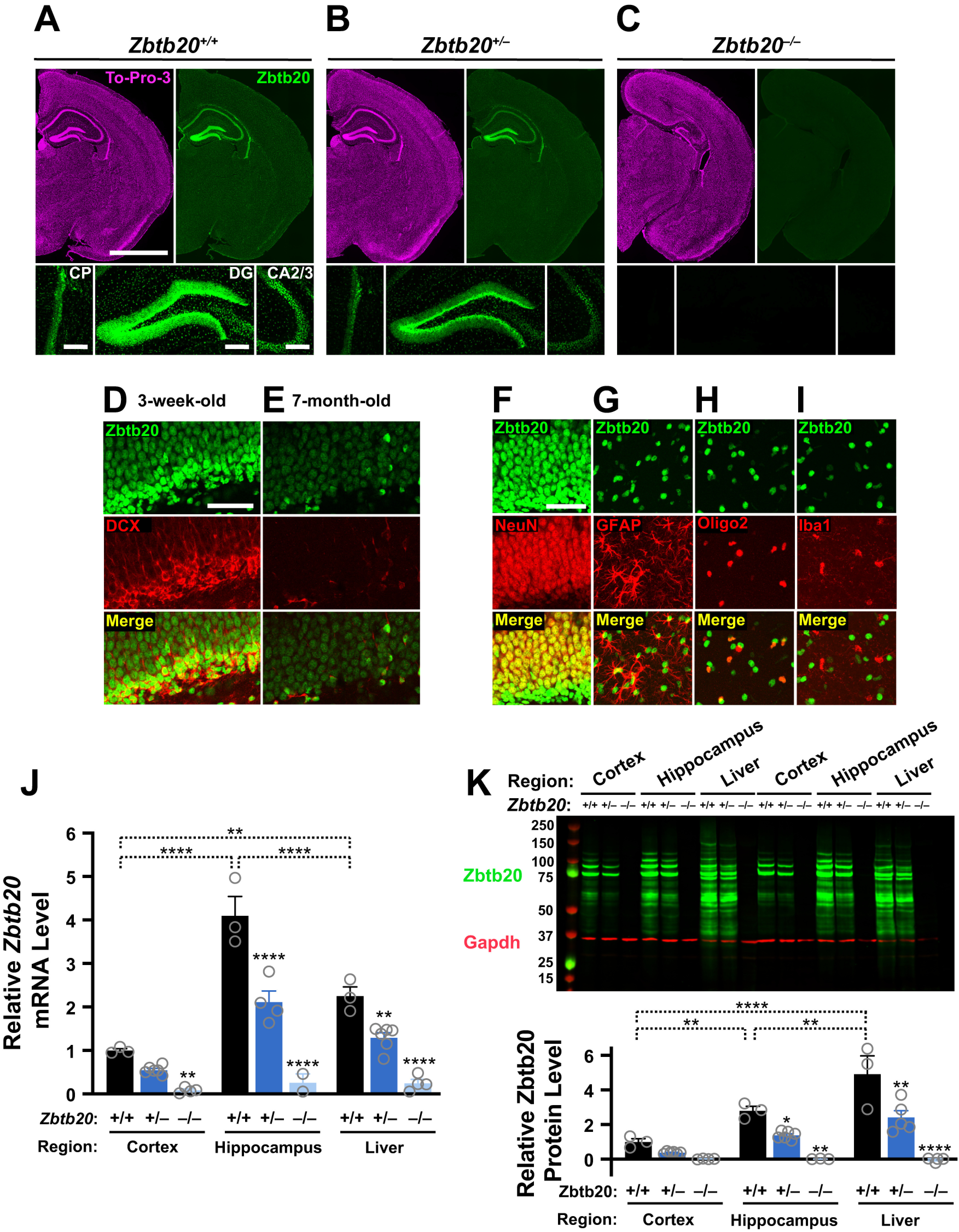
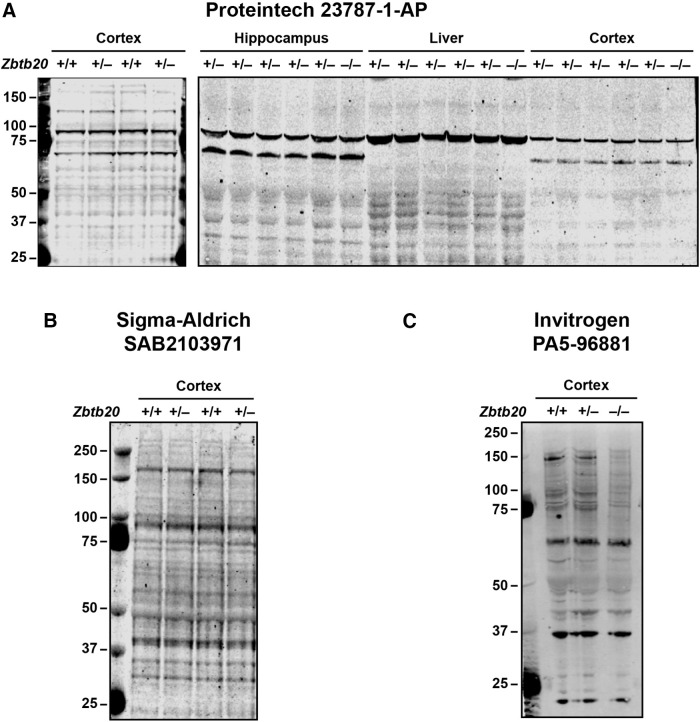
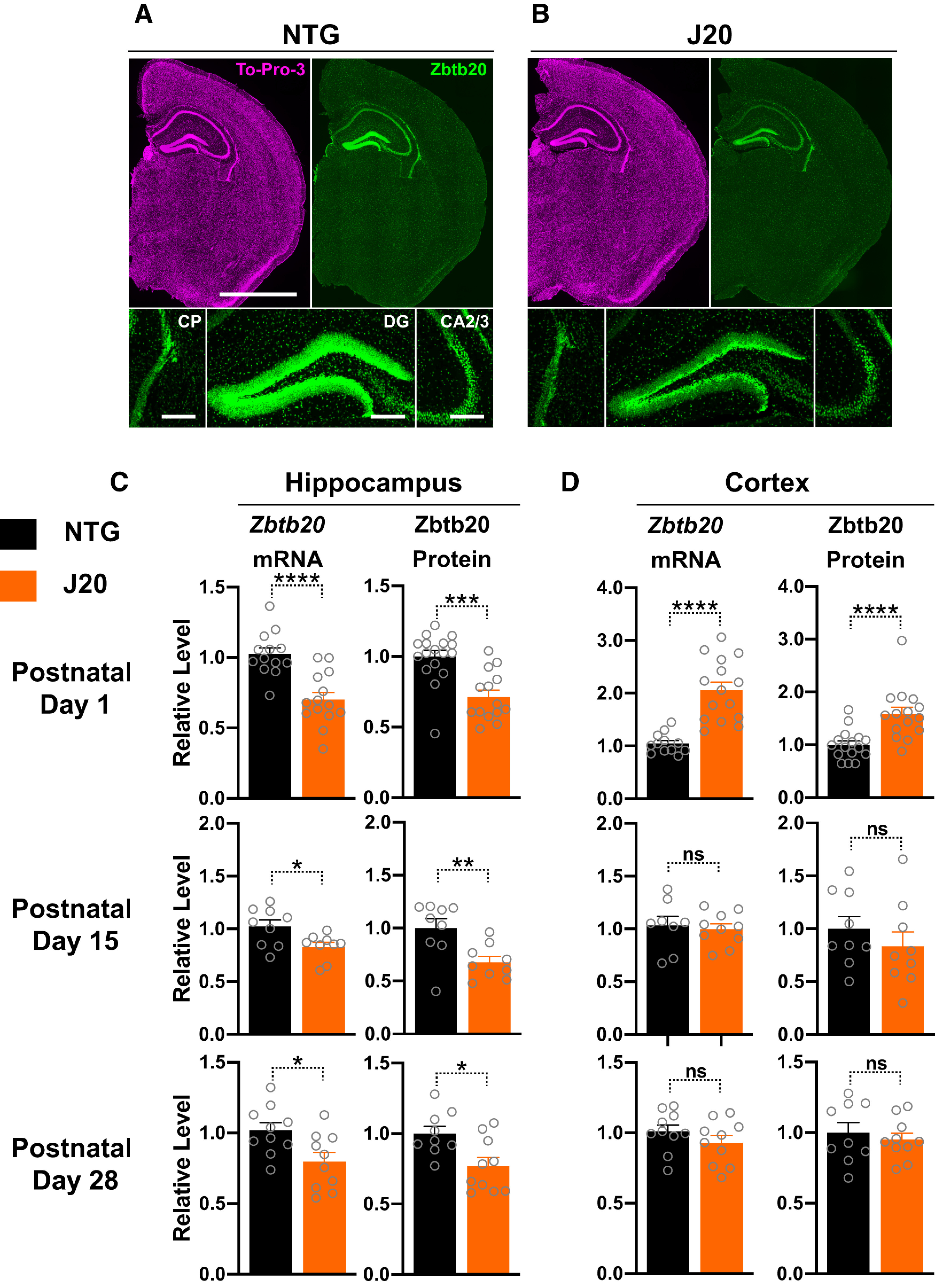
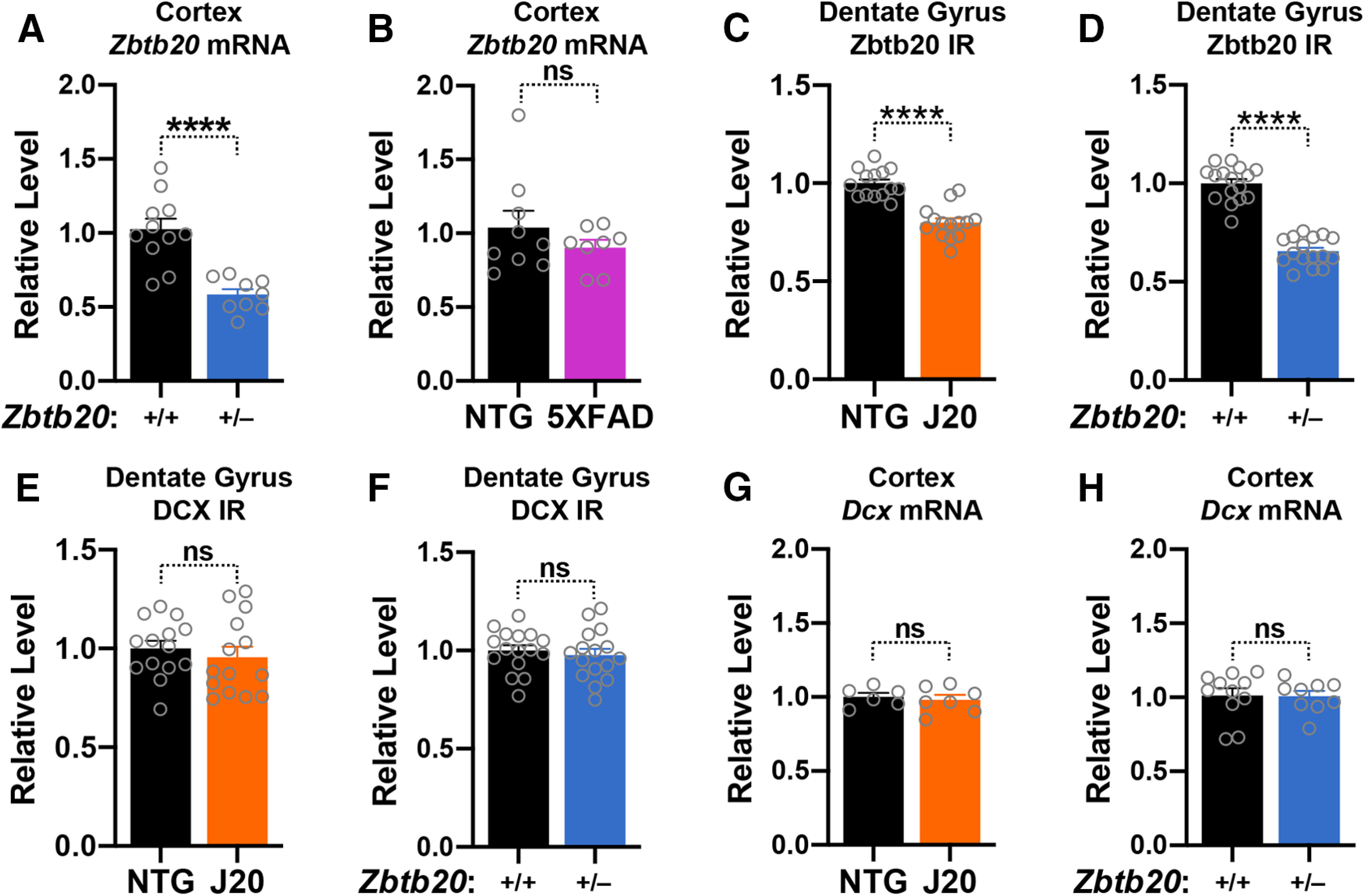
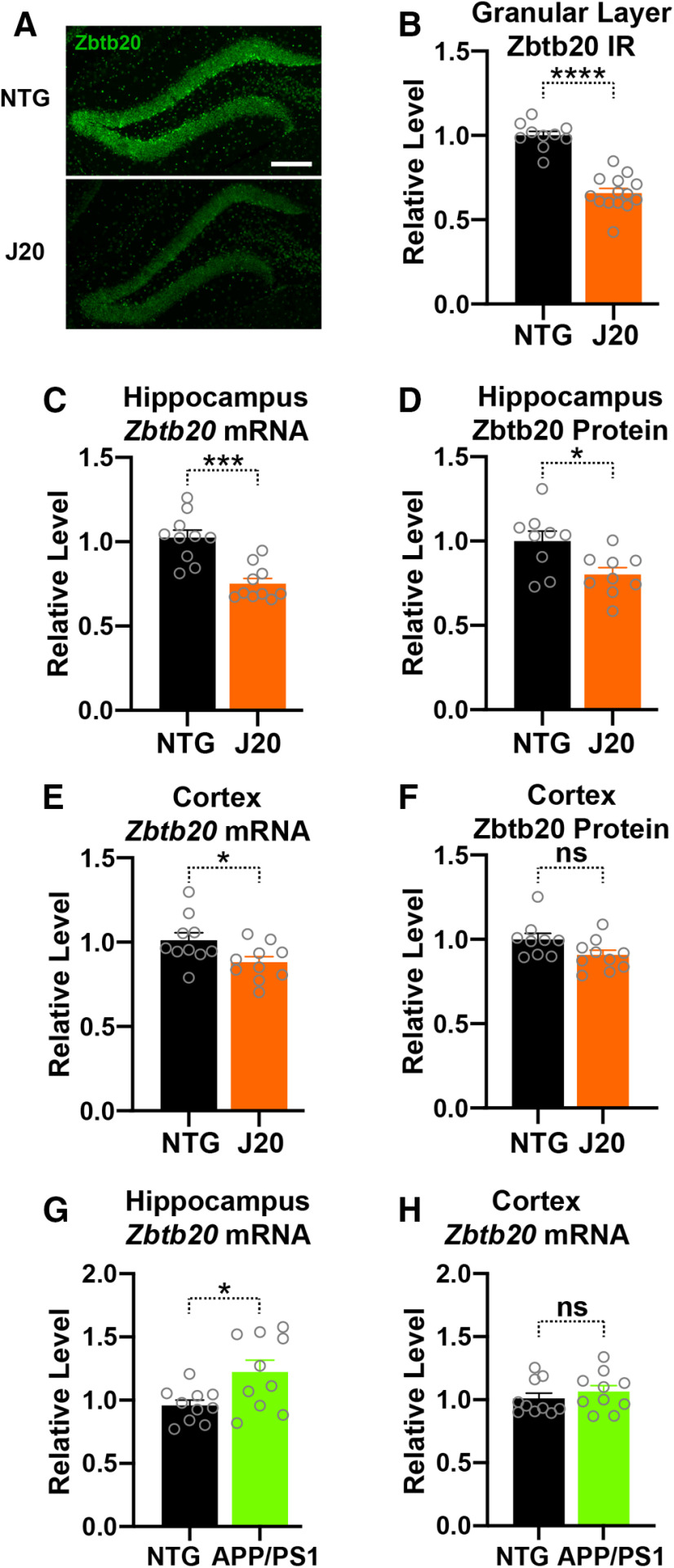
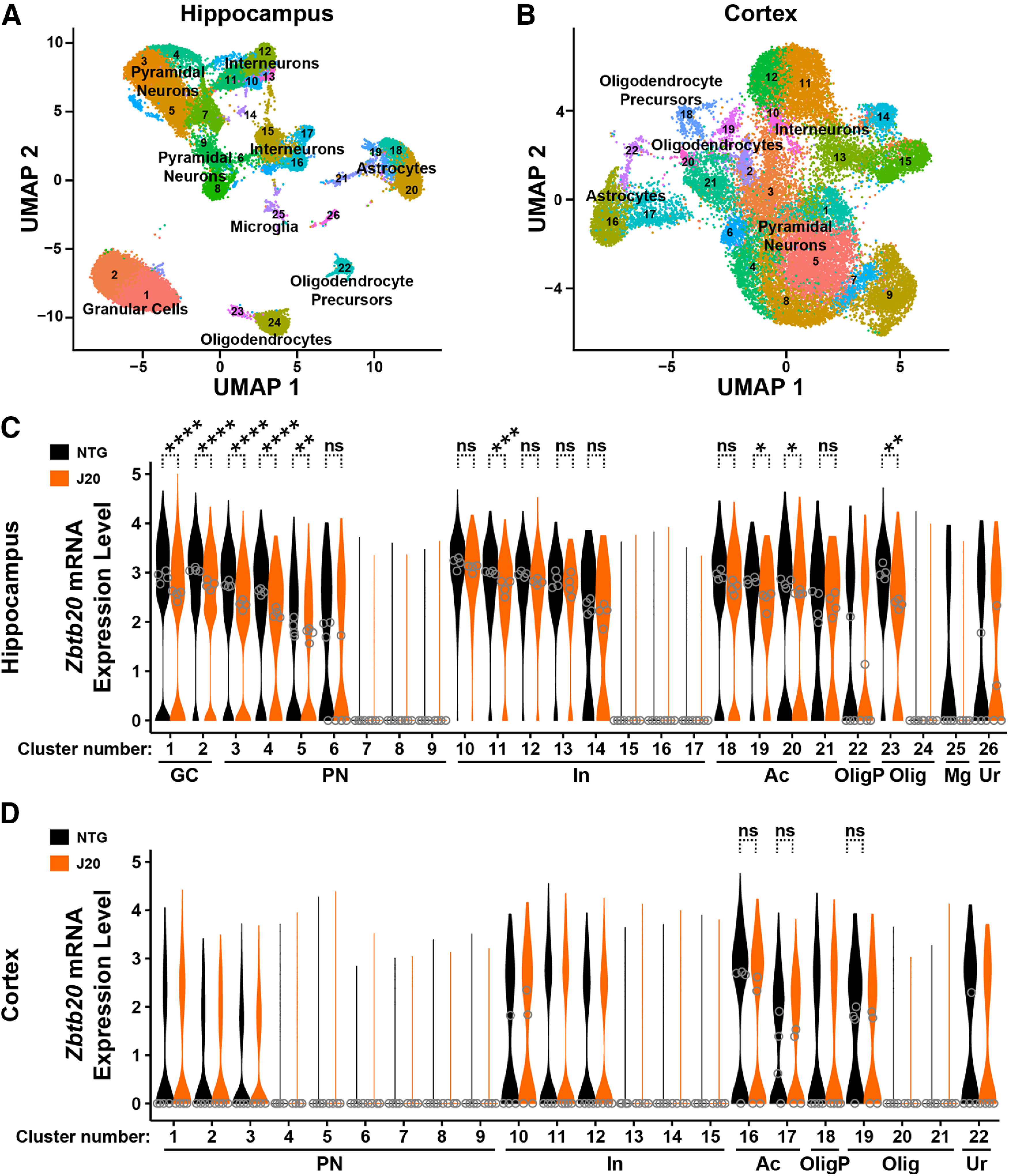
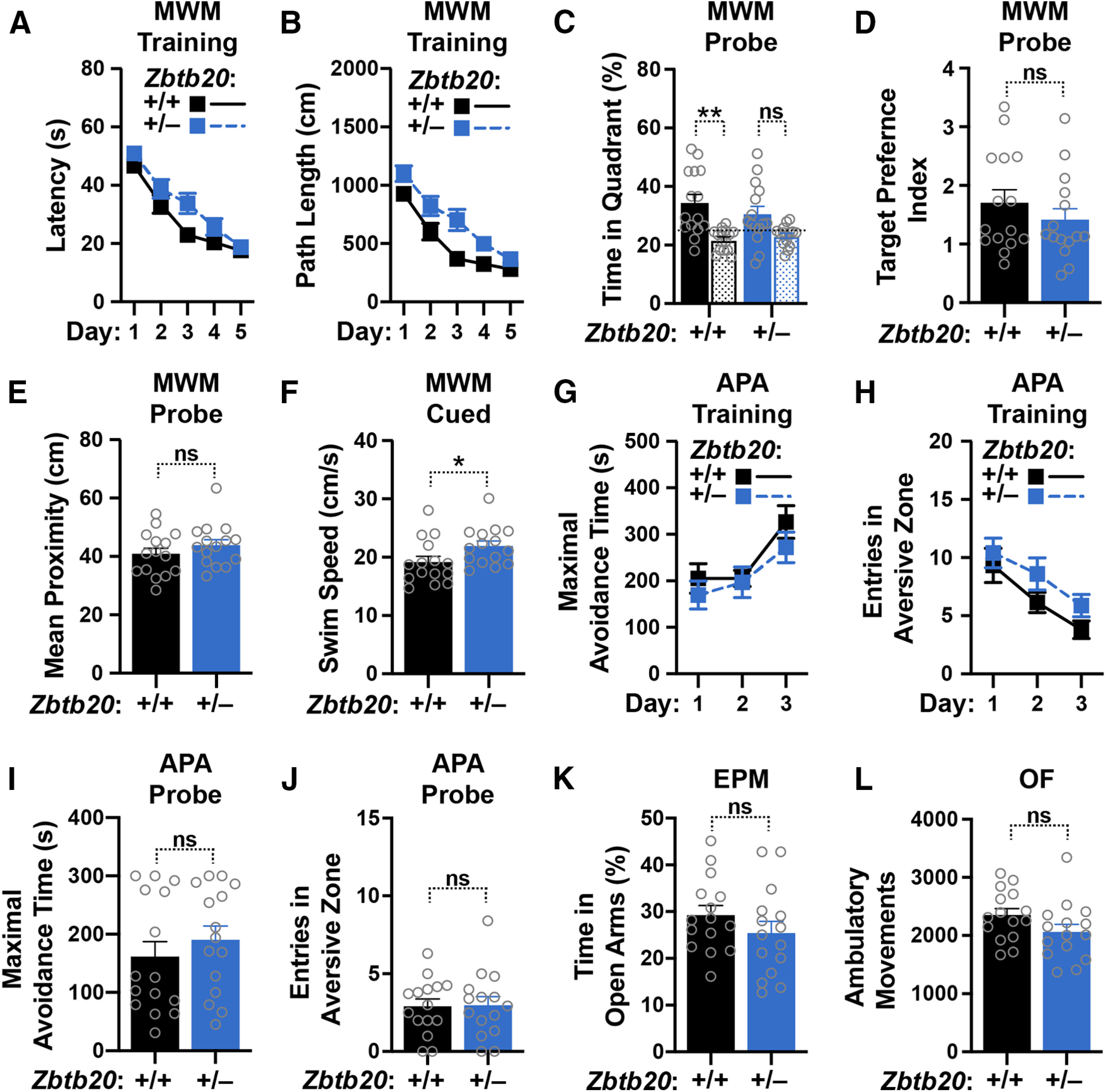
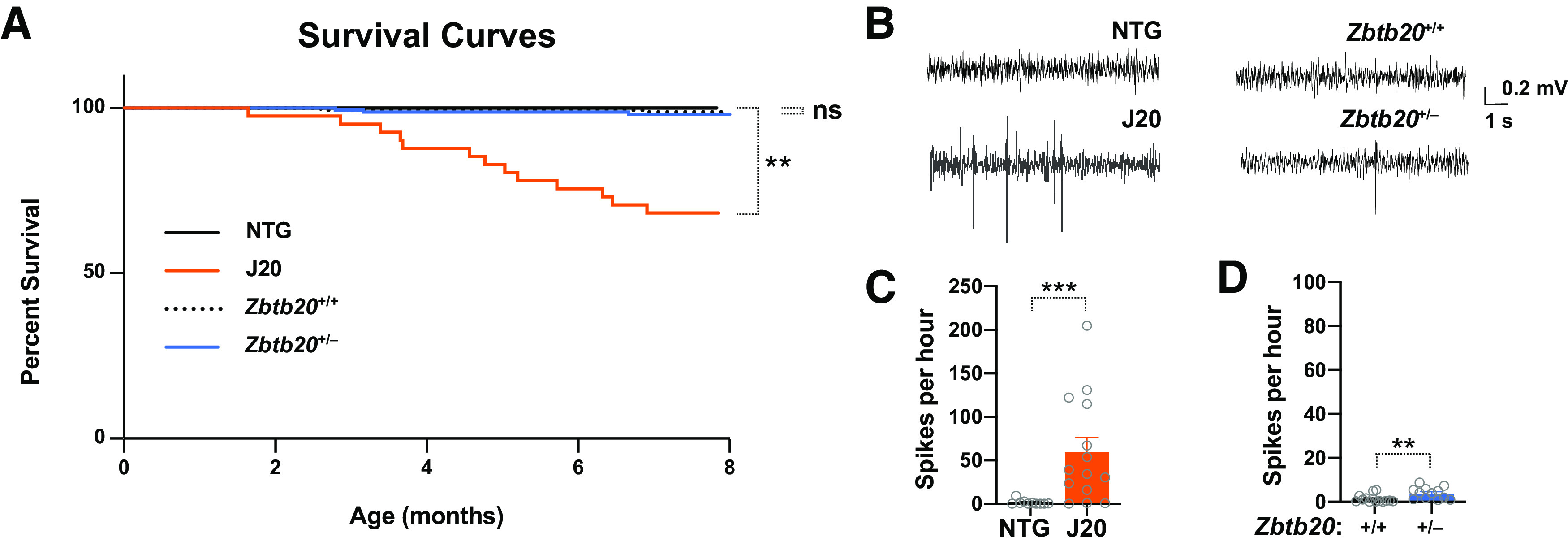
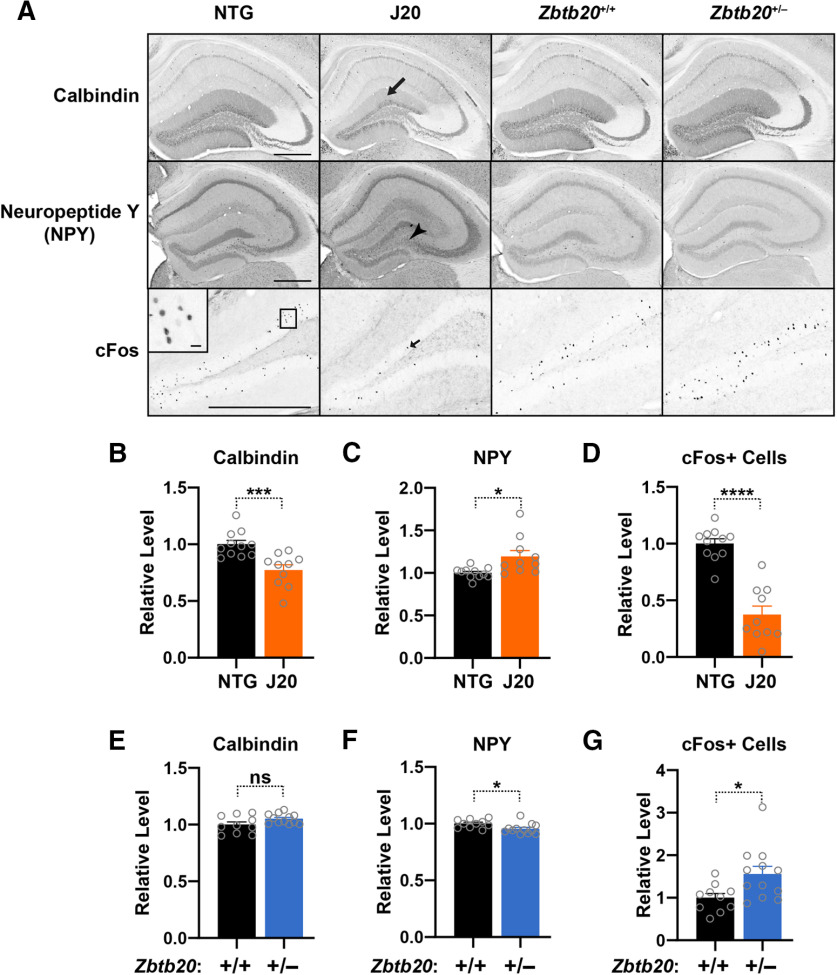
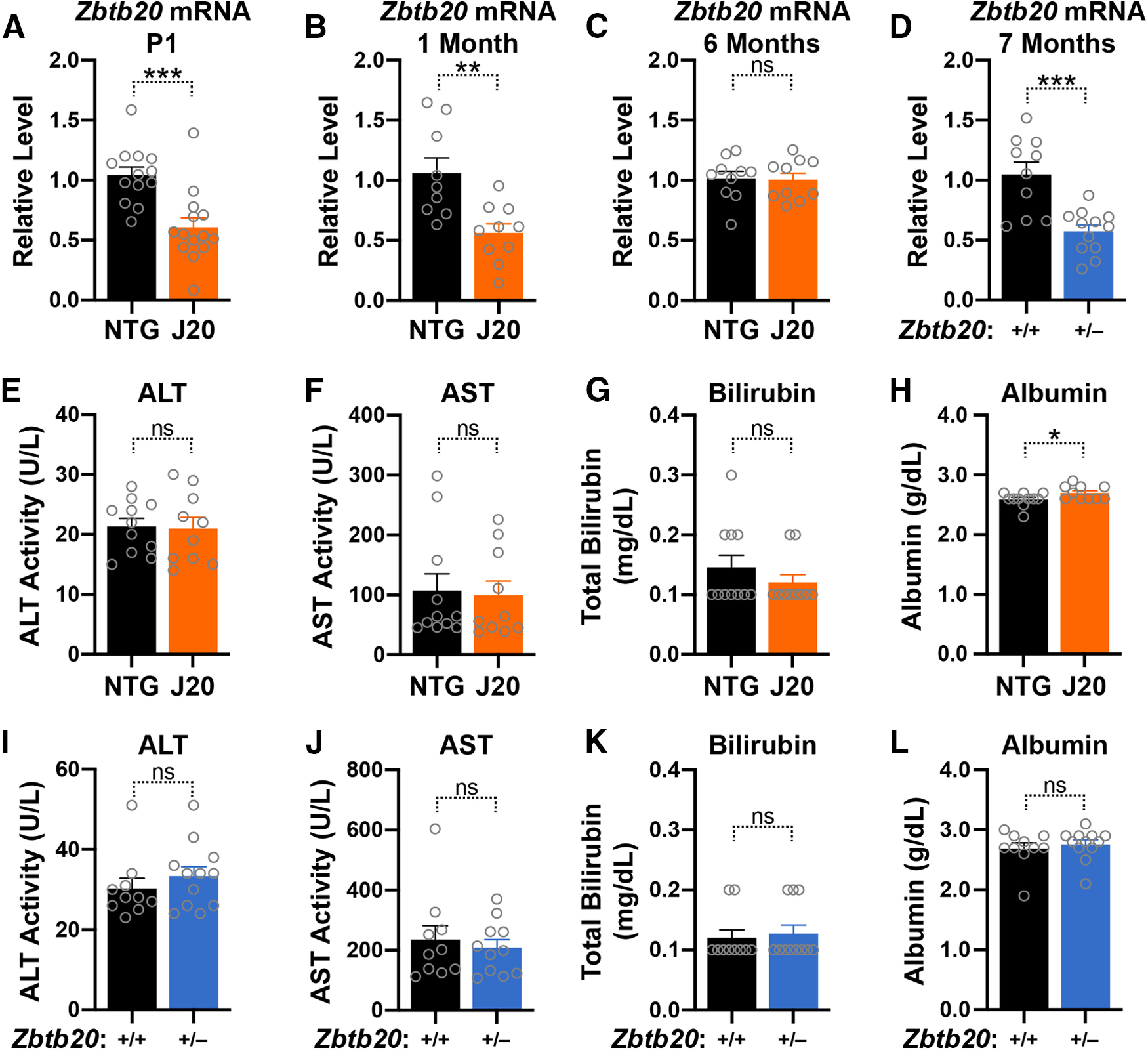
Diverse gene products contribute to the pathogenesis of Alzheimer's disease (AD). Experimental models have helped elucidate their mechanisms and impact on brain functions. Human amyloid precursor protein (hAPP) transgenic mice from line J20 (hAPP-J20 mice) are widely used to simulate key aspects of AD. However, they also carry an insertional mutation in noncoding sequence of one Zbtb20 allele, a gene involved in neural development. We demonstrate that heterozygous hAPP-J20 mice have reduced Zbtb20 expression in some AD-relevant brain regions, but not others, and that Zbtb20 levels are higher in hAPP-J20 mice than heterozygous Zbtb20 knock-out (Zbtb20+/-) mice. Whereas hAPP-J20 mice have premature mortality, severe deficits in learning and memory, other behavioral alterations, and prominent nonconvulsive epileptiform activity, Zbtb20+/- mice do not. Thus, the insertional mutation in hAPP-J20 mice does not ablate the affected Zbtb20 allele and is unlikely to account for the AD-like phenotype of this model.
Keywords: Alzheimer’s disease; Zbtb20; amyloid precursor protein; behavior; epilepsy; mouse model.
Copyright © 2021 Gulbranson et al.
Figures
References
-
- Cheng IH, Scearce-Levie K, Legleiter J, Palop JJ, Gerstein H, Bien-Ly N, Puoliväli J, Lesné S, Ashe KH, Muchowski PJ, Mucke L (2007) Accelerating amyloid-beta fibrillization reduces oligomer levels and functional deficits in Alzheimer disease mouse models. J Biol Chem 282:23818–23828. 10.1074/jbc.M701078200 - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases