

Smc5/6 functions with Sgs1-Top3-Rmi1 to complete chromosome replication at natural pause sites
- PMID: 33833229
- PMCID: PMC8032827
- DOI: 10.1038/s41467-021-22217-w
Smc5/6 functions with Sgs1-Top3-Rmi1 to complete chromosome replication at natural pause sites
Abstract
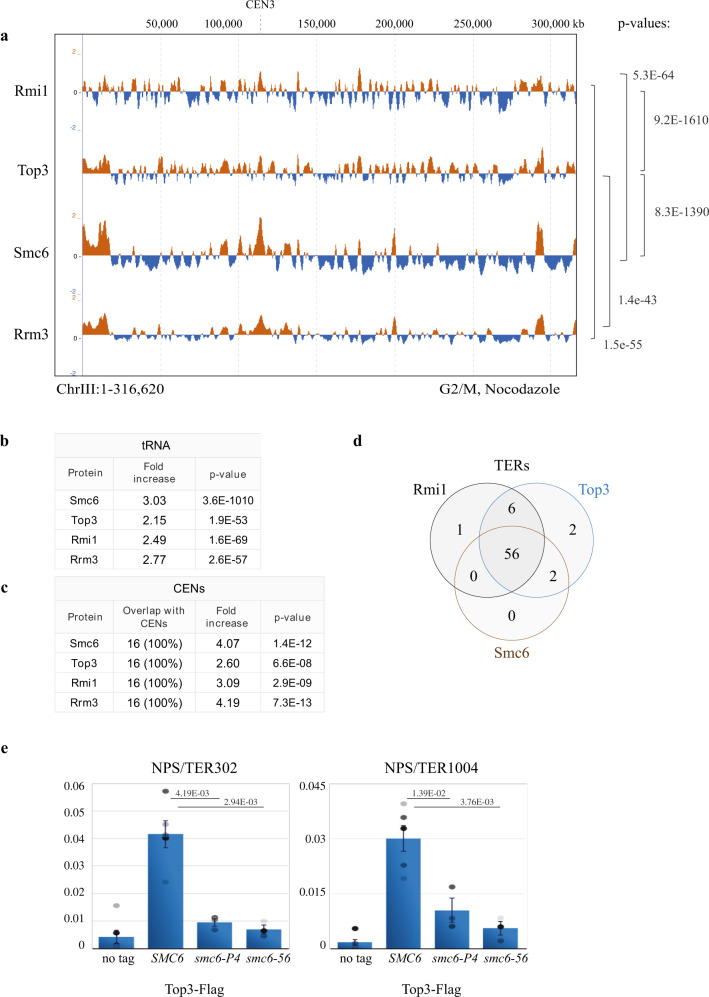
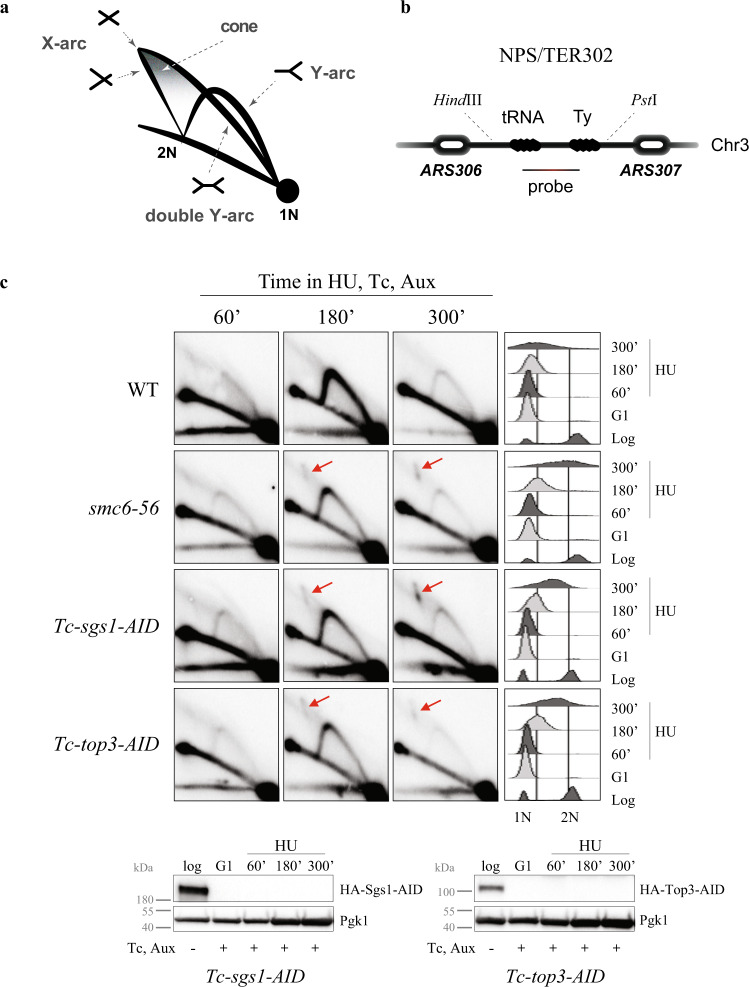
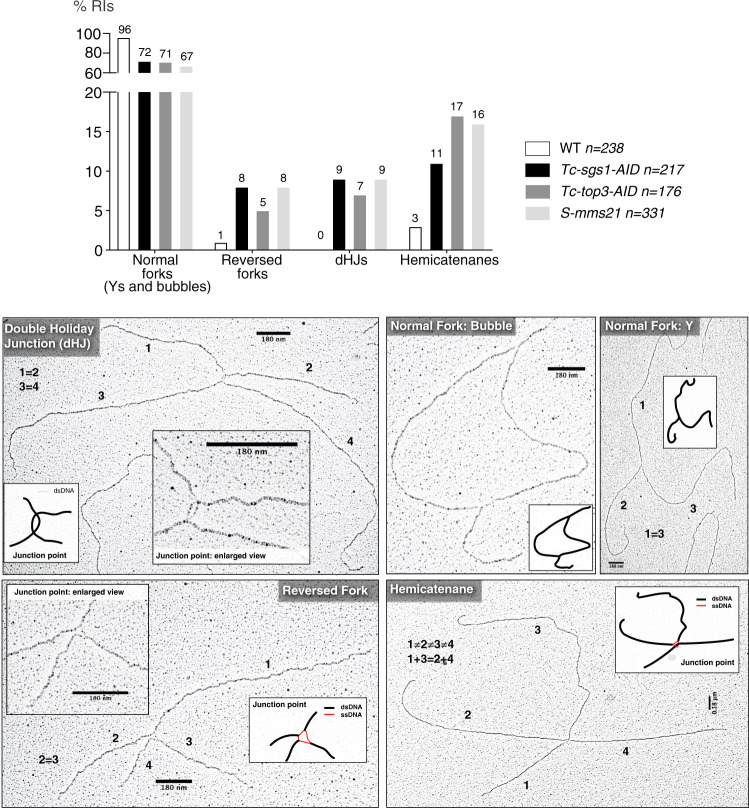
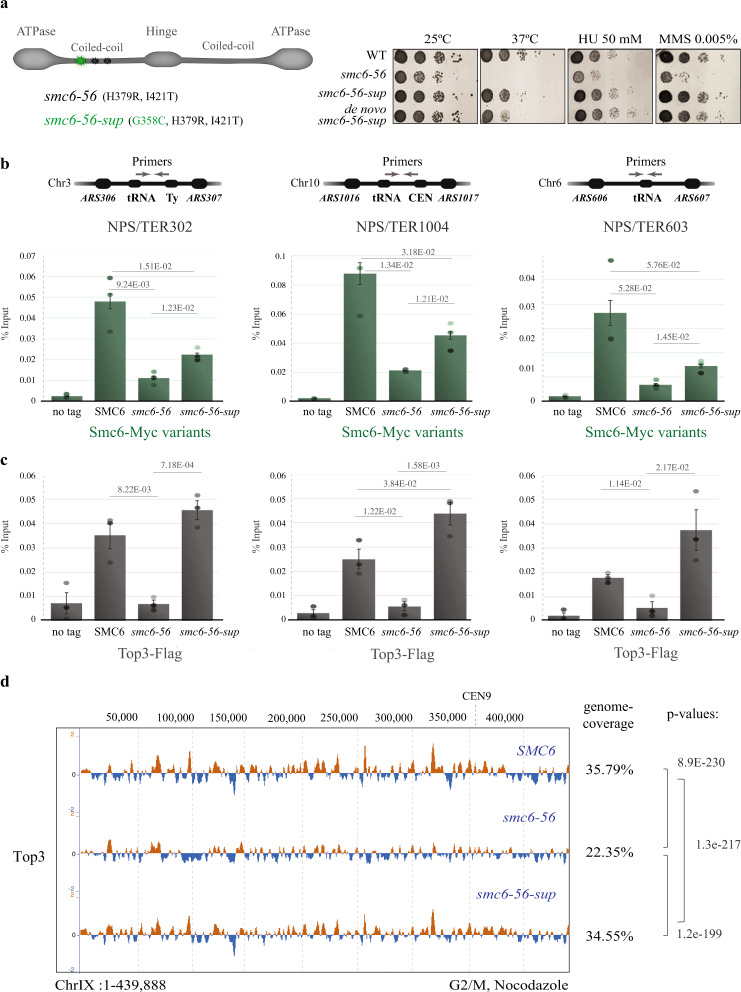
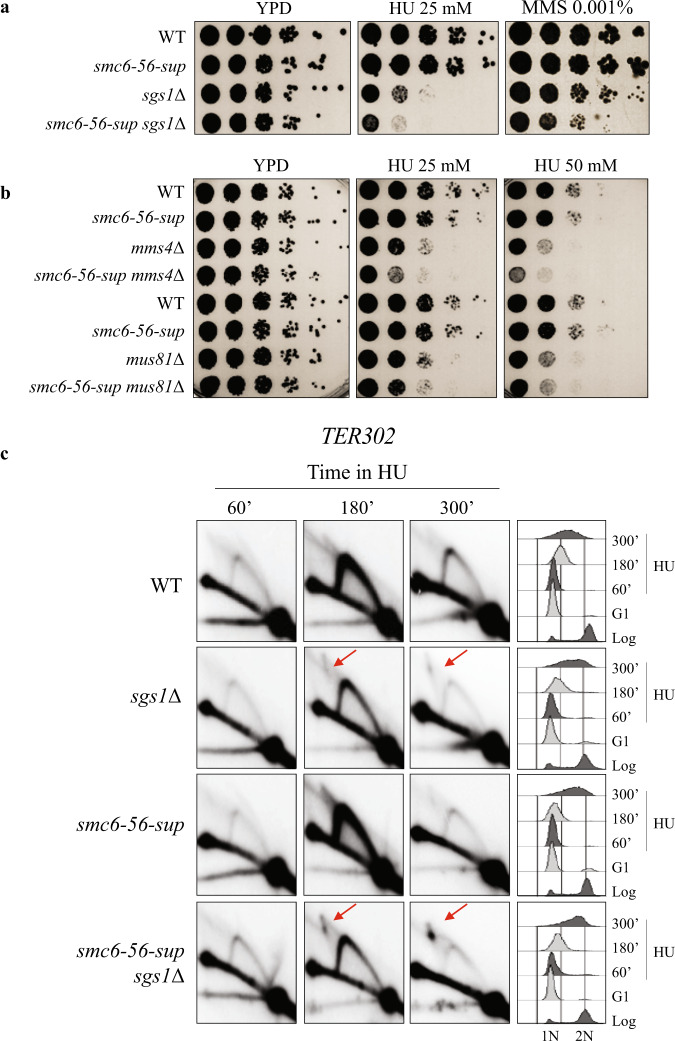
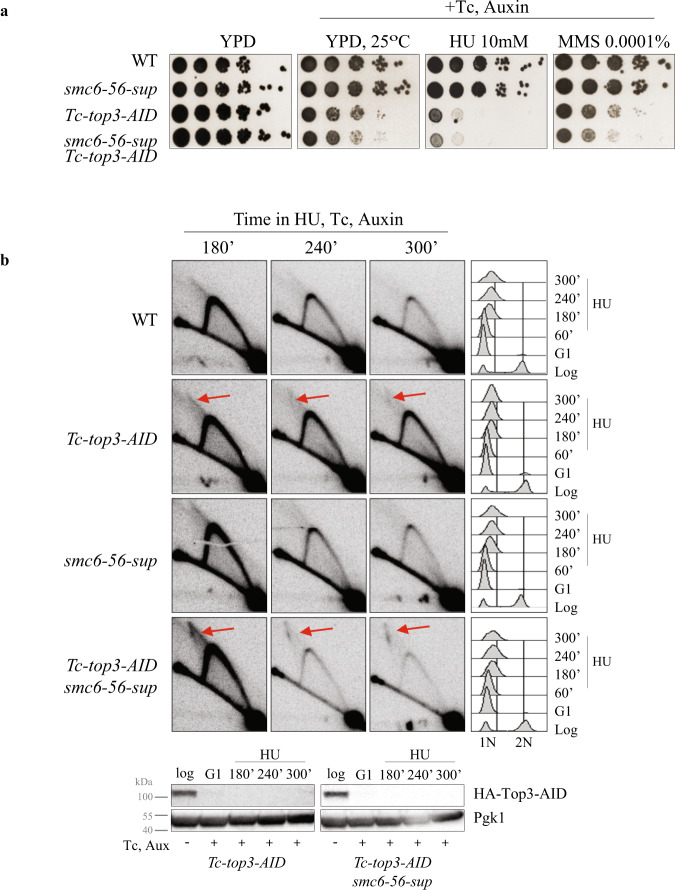
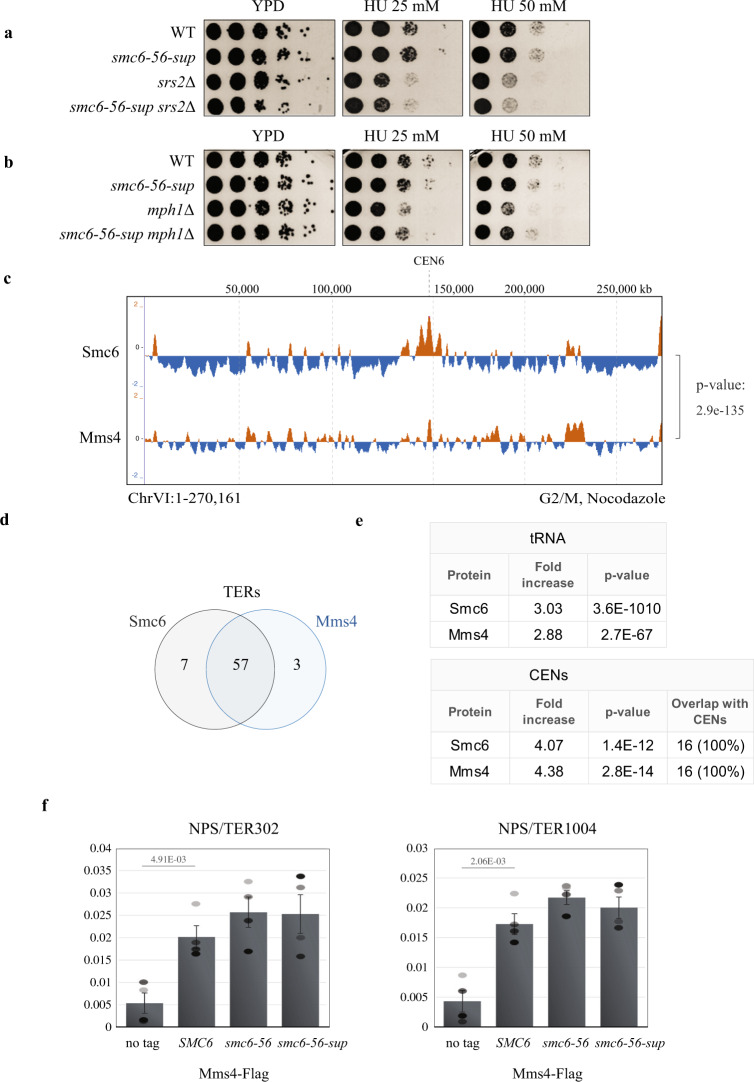
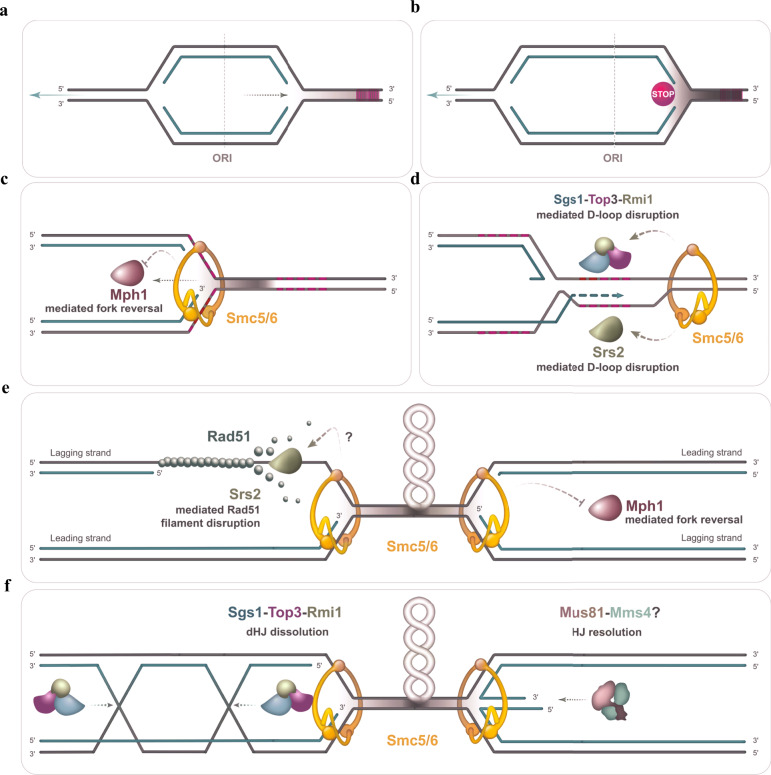
Smc5/6 is essential for genome structural integrity by yet unknown mechanisms. Here we find that Smc5/6 co-localizes with the DNA crossed-strand processing complex Sgs1-Top3-Rmi1 (STR) at genomic regions known as natural pausing sites (NPSs) where it facilitates Top3 retention. Individual depletions of STR subunits and Smc5/6 cause similar accumulation of joint molecules (JMs) composed of reversed forks, double Holliday Junctions and hemicatenanes, indicative of Smc5/6 regulating Sgs1 and Top3 DNA processing activities. We isolate an intra-allelic suppressor of smc6-56 proficient in Top3 retention but affected in pathways that act complementarily with Sgs1 and Top3 to resolve JMs arising at replication termination. Upon replication stress, the smc6-56 suppressor requires STR and Mus81-Mms4 functions for recovery, but not Srs2 and Mph1 helicases that prevent maturation of recombination intermediates. Thus, Smc5/6 functions jointly with Top3 and STR to mediate replication completion and influences the function of other DNA crossed-strand processing enzymes at NPSs.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- West, S. C. et al. Resolution of recombination intermediates: mechanisms and regulation. Cold Spring Harbor Symposia Quant. Biol.80, 103–109 (2015). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
