

Hypothesis and Theory: Roles of Arginine Methylation in C9orf72-Mediated ALS and FTD
- PMID: 33833668
- PMCID: PMC8021787
- DOI: 10.3389/fncel.2021.633668
Hypothesis and Theory: Roles of Arginine Methylation in C9orf72-Mediated ALS and FTD
Abstract
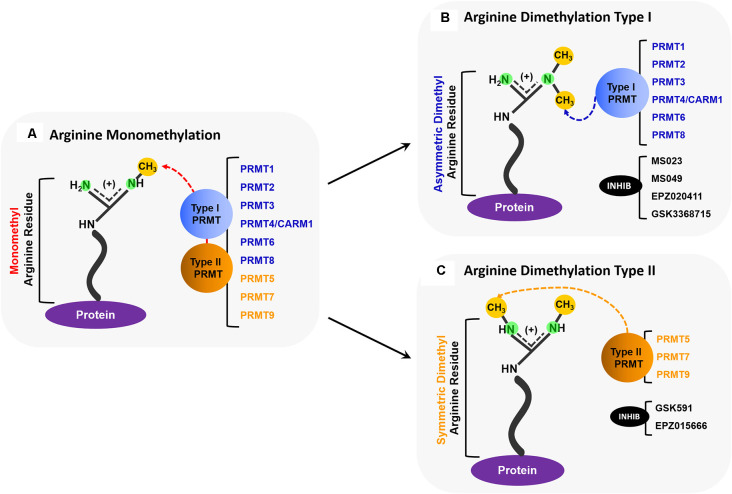
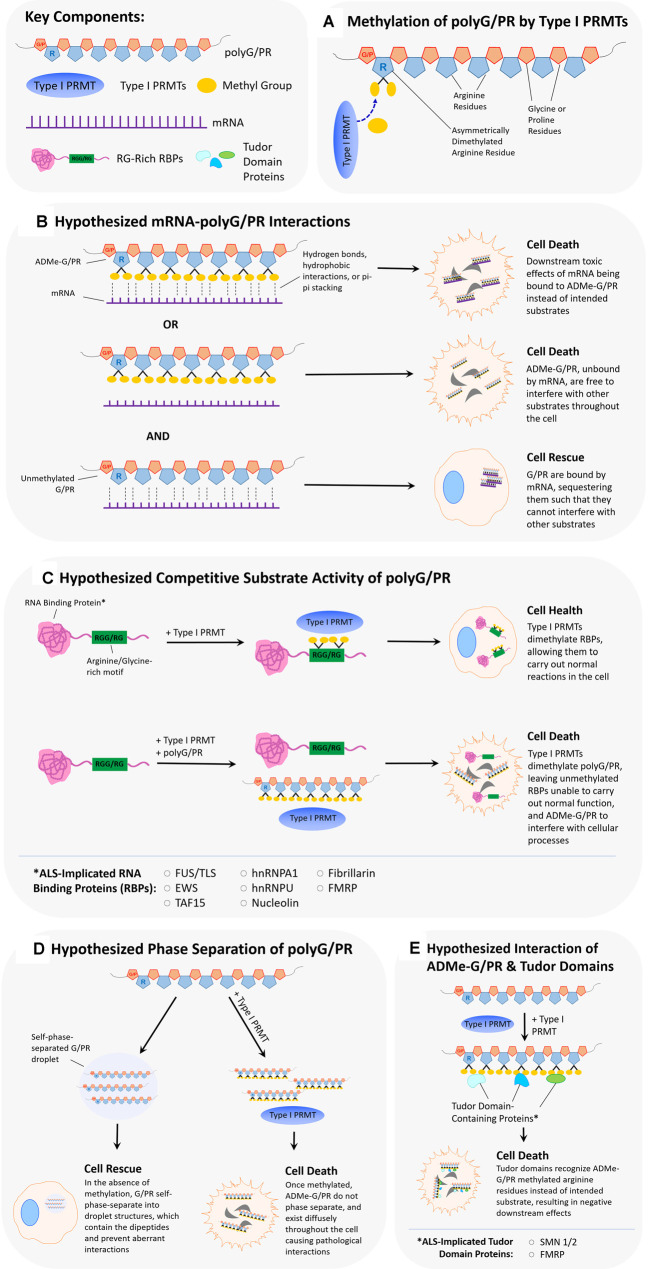
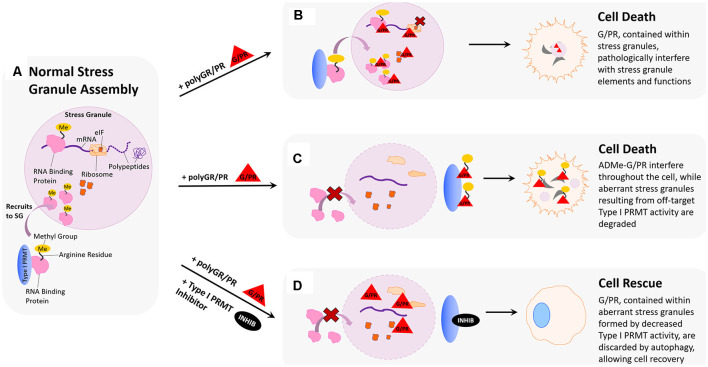
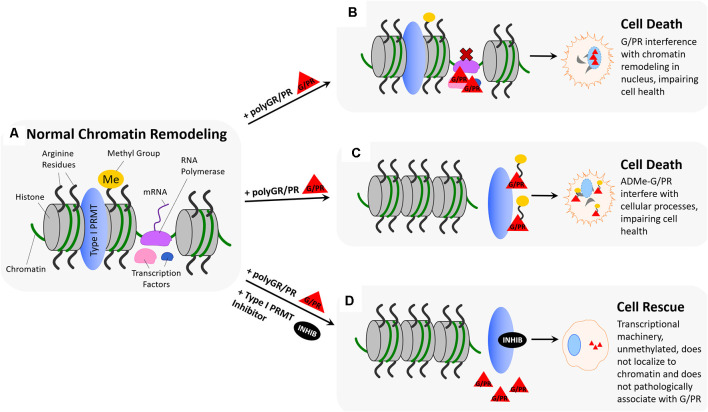
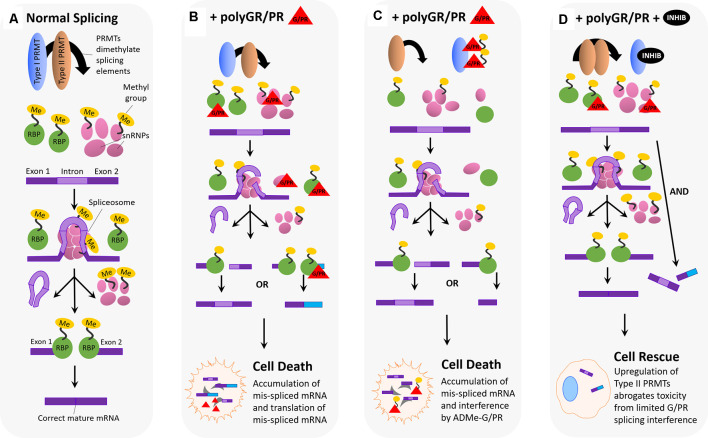
Hexanucleotide repeat expansion (G4C2n) mutations in the gene C9ORF72 account for approximately 30% of familial cases of amyotrophic lateral sclerosis (ALS) and frontotemporal dementia (FTD), as well as approximately 7% of sporadic cases of ALS. G4C2n mutations are known to result in the production of five species of dipeptide repeat proteins (DRPs) through non-canonical translation processes. Arginine-enriched dipeptide repeat proteins, glycine-arginine (polyGR), and proline-arginine (polyPR) have been demonstrated to be cytotoxic and deleterious in multiple experimental systems. Recently, we and others have implicated methylation of polyGR/polyPR arginine residues in disease processes related to G4C2n mutation-mediated neurodegeneration. We previously reported that inhibition of asymmetric dimethylation (ADMe) of arginine residues is protective in cell-based models of polyGR/polyPR cytotoxicity. These results are consistent with the idea that PRMT-mediated arginine methylation in the context of polyGR/polyPR exposure is harmful. However, it remains unclear why. Here we discuss the influence of arginine methylation on diverse cellular processes including liquid-liquid phase separation, chromatin remodeling, transcription, RNA processing, and RNA-binding protein localization, and we consider how methylation of polyGR/polyPR may disrupt processes essential for normal cellular function and survival.
Keywords: C9ORF72 ALS/FTD; PRMT inhibitor; arginine methylation; chromatin remodeling; dipeptide repeat; polyGR; polyPR; splicing.
Copyright © 2021 Gill, Premasiri and Vieira.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Reduced C9orf72 expression exacerbates polyGR toxicity in patient iPSC-derived motor neurons and a Type I protein arginine methyltransferase inhibitor reduces that toxicity.Front Cell Neurosci. 2023 Apr 17;17:1134090. doi: 10.3389/fncel.2023.1134090. eCollection 2023. Front Cell Neurosci. 2023. PMID: 37138766 Free PMC article.
-
Type I PRMT Inhibition Protects Against C9ORF72 Arginine-Rich Dipeptide Repeat Toxicity.Front Pharmacol. 2020 Sep 8;11:569661. doi: 10.3389/fphar.2020.569661. eCollection 2020. Front Pharmacol. 2020. PMID: 33013410 Free PMC article.
-
C9orf72 proline-arginine dipeptide repeats disrupt the proteasome and perturb proteolytic activities.J Neuropathol Exp Neurol. 2023 Oct 20;82(11):901-910. doi: 10.1093/jnen/nlad078. J Neuropathol Exp Neurol. 2023. PMID: 37791472 Free PMC article.
-
Mechanisms of Immune Activation by c9orf72-Expansions in Amyotrophic Lateral Sclerosis and Frontotemporal Dementia.Front Neurosci. 2019 Dec 10;13:1298. doi: 10.3389/fnins.2019.01298. eCollection 2019. Front Neurosci. 2019. PMID: 31920478 Free PMC article. Review.
-
Molecular Mechanisms of Neurodegeneration Related to C9orf72 Hexanucleotide Repeat Expansion.Behav Neurol. 2019 Jan 15;2019:2909168. doi: 10.1155/2019/2909168. eCollection 2019. Behav Neurol. 2019. PMID: 30774737 Free PMC article. Review.
Cited by
-
Disruption of the white matter structural network and its correlation with baseline progression rate in patients with sporadic amyotrophic lateral sclerosis.Transl Neurodegener. 2021 Sep 13;10(1):35. doi: 10.1186/s40035-021-00255-0. Transl Neurodegener. 2021. PMID: 34511130 Free PMC article.
-
Genome-Wide DNA Methylation in Early-Onset-Dementia Patients Brain Tissue and Lymphoblastoid Cell Lines.Int J Mol Sci. 2024 May 16;25(10):5445. doi: 10.3390/ijms25105445. Int J Mol Sci. 2024. PMID: 38791483 Free PMC article.
-
Reduced C9orf72 expression exacerbates polyGR toxicity in patient iPSC-derived motor neurons and a Type I protein arginine methyltransferase inhibitor reduces that toxicity.Front Cell Neurosci. 2023 Apr 17;17:1134090. doi: 10.3389/fncel.2023.1134090. eCollection 2023. Front Cell Neurosci. 2023. PMID: 37138766 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous