

Interactive Effects of Mycorrhizae, Soil Phosphorus, and Light on Growth and Induction and Priming of Defense in Plantago lanceolata
- PMID: 33833771
- PMCID: PMC8021950
- DOI: 10.3389/fpls.2021.647372
Interactive Effects of Mycorrhizae, Soil Phosphorus, and Light on Growth and Induction and Priming of Defense in Plantago lanceolata
Abstract
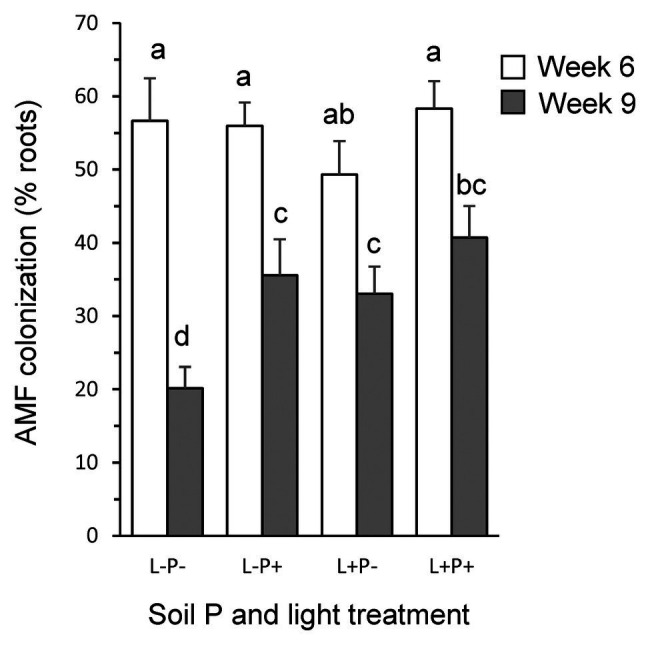
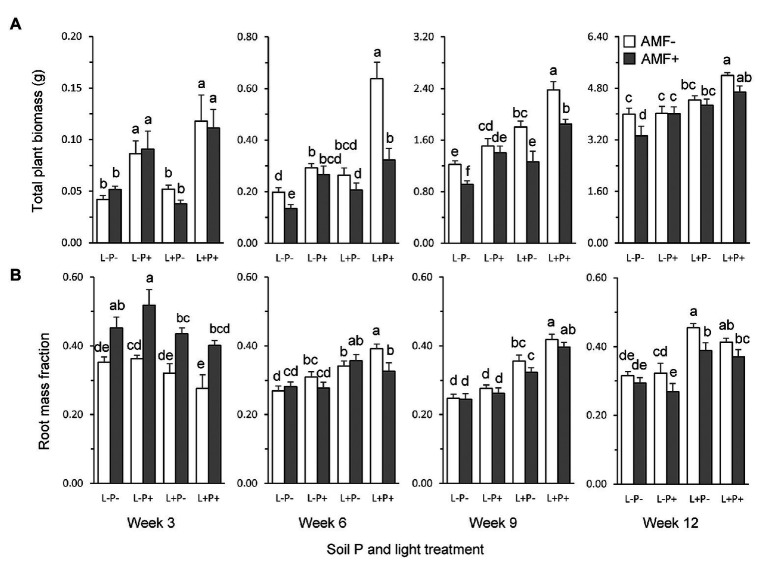
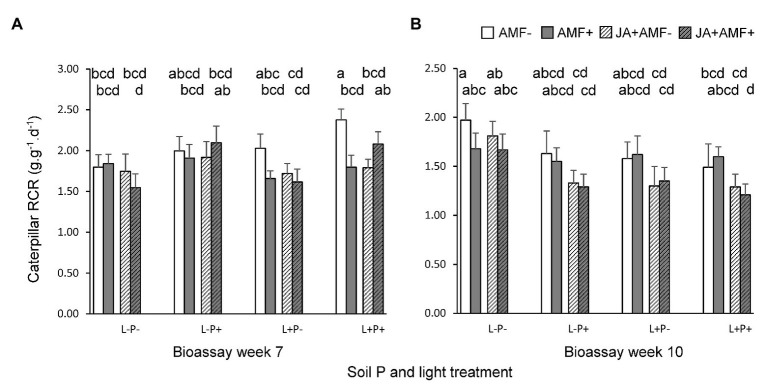
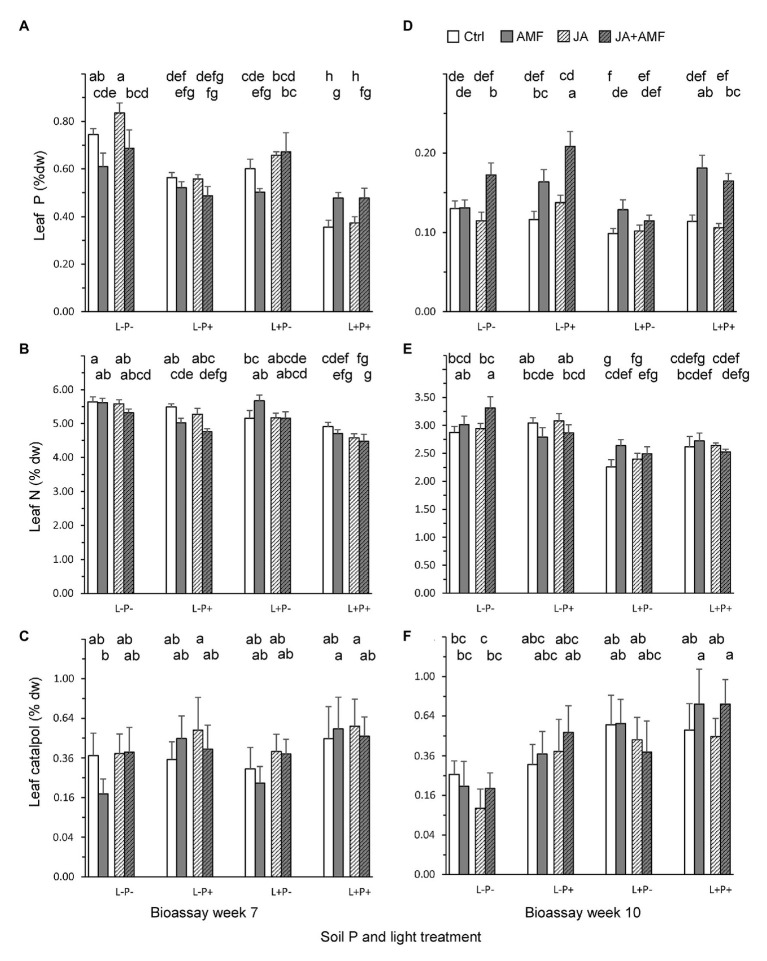
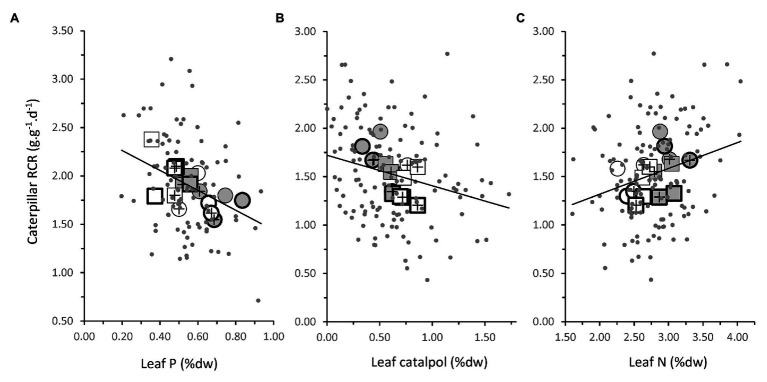
Increasing demands to reduce fertilizer and pesticide input in agriculture has triggered interest in arbuscular mycorrhizal fungi (AMF) that can enhance plant growth and confer mycorrhiza-induced resistance (MIR). MIR can be based on a variety of mechanisms, including induction of defense compounds, and sensitization of the plant's immune system (priming) for enhanced defense against later arriving pests or pathogens signaled through jasmonic acid (JA). However, growth and resistance benefits of AMF highly depend on environmental conditions. Low soil P and non-limiting light conditions are expected to enhance MIR, as these conditions favor AMF colonization and because of observed positive cross-talk between the plant's phosphate starvation response (PSR) and JA-dependent immunity. We therefore tested growth and resistance benefits of the AMF Funneliformis mosseae in Plantago lanceolata plants grown under different levels of soil P and light intensity. Resistance benefits were assessed in bioassays with the leaf chewing herbivore Mamestra brassicae. Half of the plants were induced by jasmonic acid prior to the bioassays to specifically test whether AMF primed plants for JA-signaled defense under different abiotic conditions. AMF reduced biomass production but contrary to prediction, this reduction was not strongest under conditions considered least optimal for carbon-for-nutrient trade (low light, high soil P). JA application induced resistance to M. brassicae, but its extent was independent of soil P and light conditions. Strikingly, in younger plants, JA-induced resistance was annulled by AMF under high resource conditions (high soil P, ample light), indicating that AMF did not prime but repressed JA-induced defense responses. In older plants, low soil P and light enhanced susceptibility to M. brassicae due to enhanced leaf nitrogen levels and reduced leaf levels of the defense metabolite catalpol. By contrast, in younger plants, low soil P enhanced resistance. Our results highlight that defense priming by AMF is not ubiquitous and calls for studies revealing the causes of the increasingly observed repression of JA-mediated defense by AMF. Our study further shows that in our system abiotic factors are significant modulators of defense responses, but more strongly so by directly modulating leaf quality than by modulating the effects of beneficial microbes on resistance.
Keywords: Funneliformis mosseae; Mamestra brassicae; defense priming; induced systemic resistance; iridoid glycosides; mycorrhiza-induced resistance, shading; soil phosphorus.
Copyright © 2021 Qu, Wang and Biere.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Effects of arbuscular mycorrhizal fungi on plant growth and herbivore infestation depend on availability of soil water and nutrients.Front Plant Sci. 2023 Jan 26;14:1101932. doi: 10.3389/fpls.2023.1101932. eCollection 2023. Front Plant Sci. 2023. PMID: 36778709 Free PMC article.
-
Chemical defense, mycorrhizal colonization and growth responses in Plantago lanceolata L.Oecologia. 2009 Jun;160(3):433-42. doi: 10.1007/s00442-009-1312-2. Epub 2009 Mar 7. Oecologia. 2009. PMID: 19271240
-
Effects of the Timing of Herbivory on Plant Defense Induction and Insect Performance in Ribwort Plantain (Plantago lanceolata L.) Depend on Plant Mycorrhizal Status.J Chem Ecol. 2015 Nov;41(11):1006-17. doi: 10.1007/s10886-015-0644-0. Epub 2015 Nov 9. J Chem Ecol. 2015. PMID: 26552915 Free PMC article.
-
Role of Arbuscular Mycorrhizal Fungi in Regulating Growth, Enhancing Productivity, and Potentially Influencing Ecosystems under Abiotic and Biotic Stresses.Plants (Basel). 2023 Aug 29;12(17):3102. doi: 10.3390/plants12173102. Plants (Basel). 2023. PMID: 37687353 Free PMC article. Review.
-
Arbuscular Mycorrhizal Symbiosis: Plant Friend or Foe in the Fight Against Viruses?Front Microbiol. 2019 Jun 4;10:1238. doi: 10.3389/fmicb.2019.01238. eCollection 2019. Front Microbiol. 2019. PMID: 31231333 Free PMC article. Review.
Cited by
-
Mycorrhiza-Induced Resistance against Foliar Pathogens Is Uncoupled of Nutritional Effects under Different Light Intensities.J Fungi (Basel). 2021 May 21;7(6):402. doi: 10.3390/jof7060402. J Fungi (Basel). 2021. PMID: 34063889 Free PMC article.
-
Effects of arbuscular mycorrhizal fungi on plant growth and herbivore infestation depend on availability of soil water and nutrients.Front Plant Sci. 2023 Jan 26;14:1101932. doi: 10.3389/fpls.2023.1101932. eCollection 2023. Front Plant Sci. 2023. PMID: 36778709 Free PMC article.
-
Testing the joint effects of arbuscular mycorrhizal fungi and ants on insect herbivory on potato plants.Planta. 2024 Jul 30;260(3):66. doi: 10.1007/s00425-024-04492-1. Planta. 2024. PMID: 39080142 Free PMC article.
-
Effects of Light Quality on Colonization of Tomato Roots by AMF and Implications for Growth and Defense.Plants (Basel). 2022 Mar 24;11(7):861. doi: 10.3390/plants11070861. Plants (Basel). 2022. PMID: 35406841 Free PMC article.
-
Arbuscular mycorrhizal fungi influence host infection during epidemics in a wild plant pathosystem.New Phytol. 2022 Dec;236(5):1922-1935. doi: 10.1111/nph.18481. Epub 2022 Oct 7. New Phytol. 2022. PMID: 36093733 Free PMC article.
References
-
- Andrade S. A. L., Malik S., Sawaya A., Bottcher A., Mazzafera P. (2013). Association with arbuscular mycorrhizal fungi influences alkaloid synthesis and accumulation in Catharanthus roseus and Nicotiana tabacum plants. Acta Physiol. Plant. 35, 867–880. 10.1007/s11738-012-1130-8 - DOI
-
- Ayres R. L., Gange A. C., Aplin D. M. (2006). Interactions between arbuscular mycorrhizal fungi and intraspecific competition affect size, and size inequality, of Plantago lanceolata L. J. Ecol. 94, 285–294. 10.1111/j.1365-2745.2006.01103.x - DOI
-
- Barber N. A. (2013). Arbuscular mycorrhizal fungi are necessary for the induced response to herbivores by Cucumis sativus. J. Plant Ecol. 6, 171–176. 10.1093/jpe/rts026 - DOI
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources