

Structure versus function: Are new conformations of pannexin 1 yet to be resolved?
- PMID: 33835130
- PMCID: PMC8042604
- DOI: 10.1085/jgp.202012754
Structure versus function: Are new conformations of pannexin 1 yet to be resolved?
Abstract
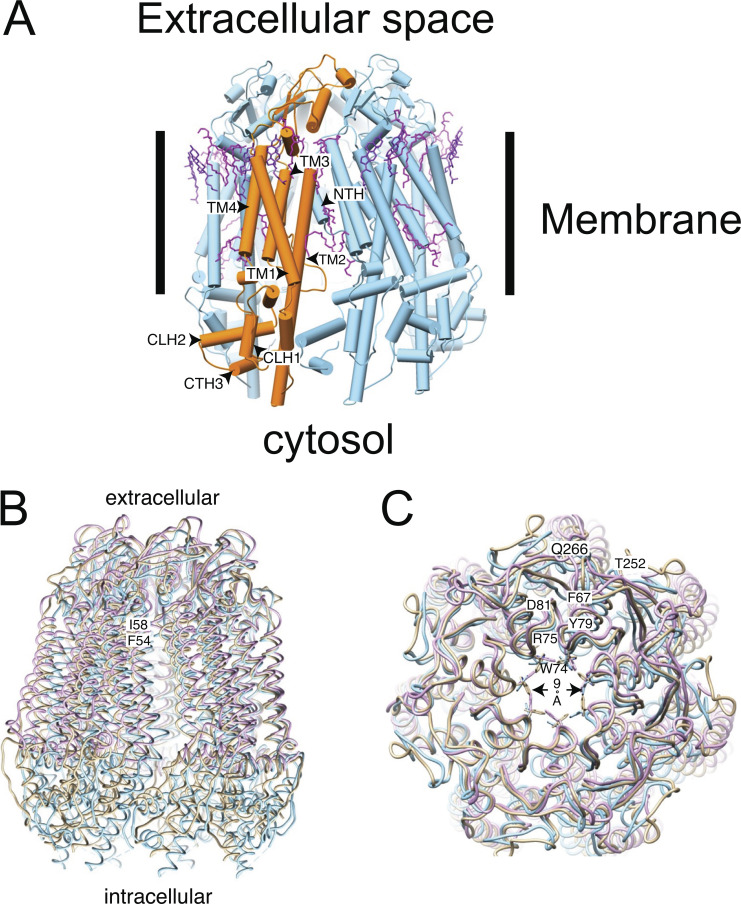
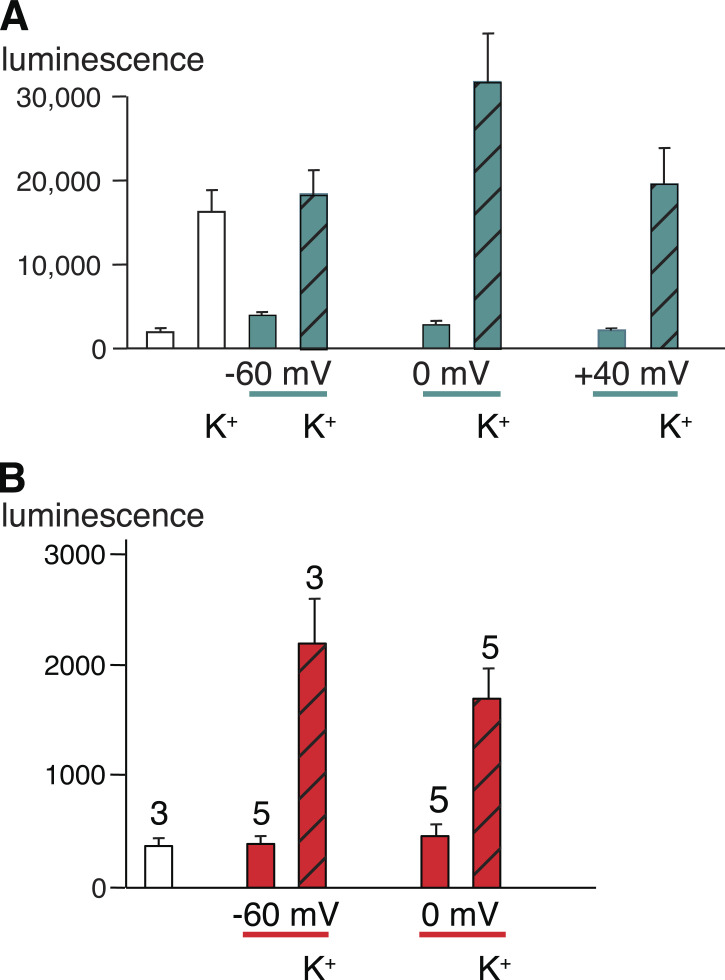
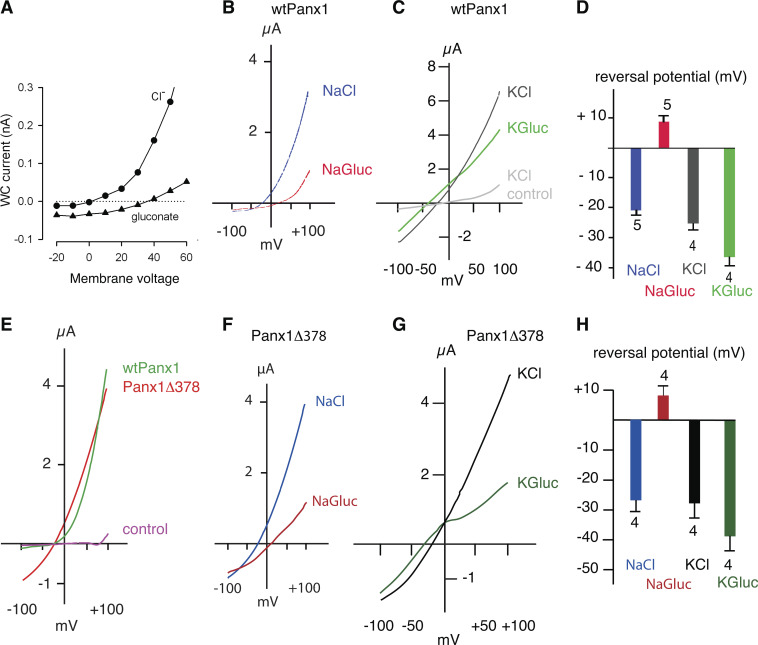
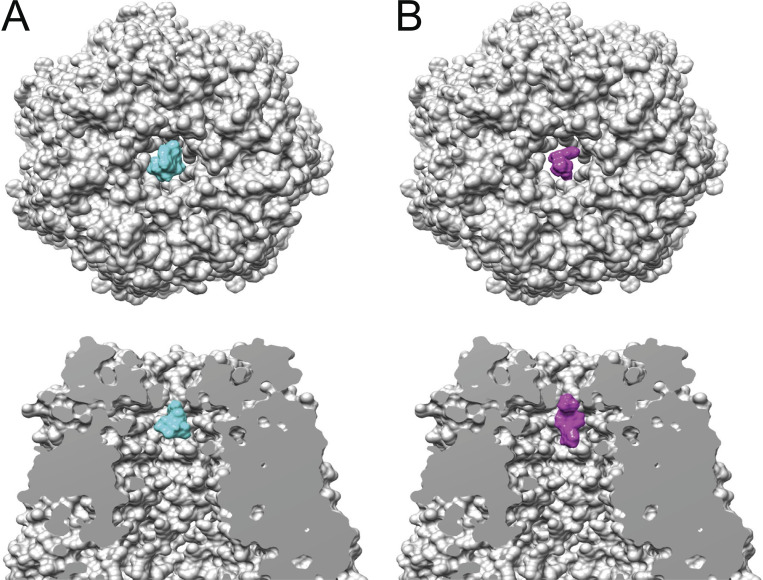
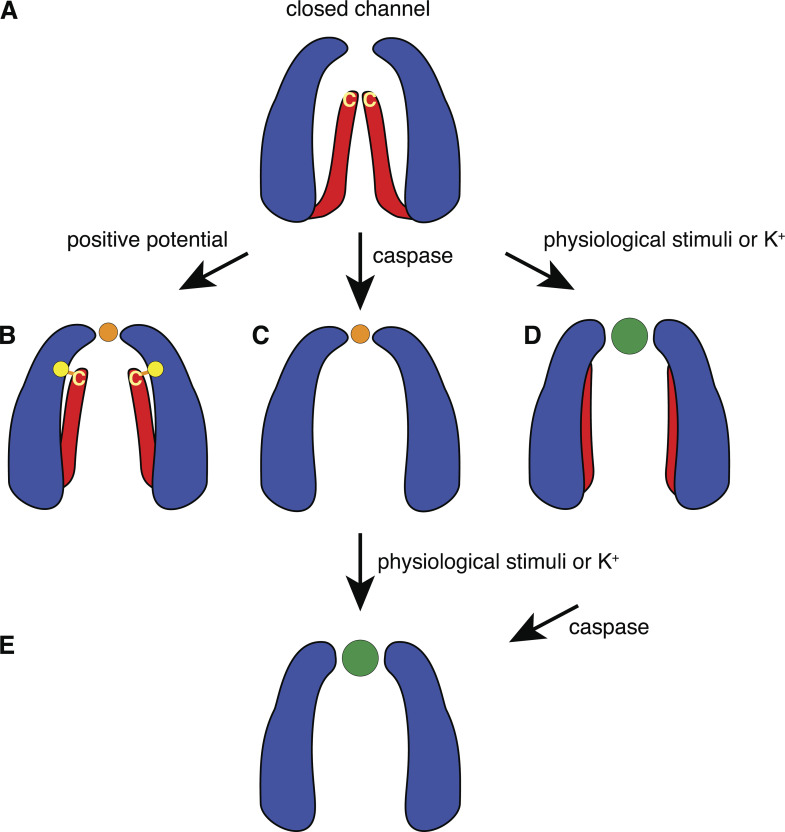
Pannexin 1 (Panx1) plays a decisive role in multiple physiological and pathological settings, including oxygen delivery to tissues, mucociliary clearance in airways, sepsis, neuropathic pain, and epilepsy. It is widely accepted that Panx1 exerts its role in the context of purinergic signaling by providing a transmembrane pathway for ATP. However, under certain conditions, Panx1 can also act as a highly selective membrane channel for chloride ions without ATP permeability. A recent flurry of publications has provided structural information about the Panx1 channel. However, while these structures are consistent with a chloride selective channel, none show a conformation with strong support for the ATP release function of Panx1. In this Viewpoint, we critically assess the existing evidence for the function and structure of the Panx1 channel and conclude that the structure corresponding to the ATP permeation pathway is yet to be determined. We also list a set of additional topics needing attention and propose ways to attain the large-pore, ATP-permeable conformation of the Panx1 channel.
© 2021 Mim et al.
Figures
Similar articles
-
Cationic control of Panx1 channel function.Am J Physiol Cell Physiol. 2018 Sep 1;315(3):C279-C289. doi: 10.1152/ajpcell.00303.2017. Epub 2018 May 2. Am J Physiol Cell Physiol. 2018. PMID: 29719168
-
ATP and large signaling metabolites flux through caspase-activated Pannexin 1 channels.Elife. 2021 Jan 7;10:e64787. doi: 10.7554/eLife.64787. Elife. 2021. PMID: 33410749 Free PMC article.
-
The Pannexin1 membrane channel: distinct conformations and functions.FEBS Lett. 2018 Oct;592(19):3201-3209. doi: 10.1002/1873-3468.13115. Epub 2018 Jun 5. FEBS Lett. 2018. PMID: 29802622 Review.
-
Pannexin1: a multifunction and multiconductance and/or permeability membrane channel.Am J Physiol Cell Physiol. 2018 Sep 1;315(3):C290-C299. doi: 10.1152/ajpcell.00302.2017. Epub 2018 May 2. Am J Physiol Cell Physiol. 2018. PMID: 29719171
-
Human Pannexin 1 channel: Insight in structure-function mechanism and its potential physiological roles.Mol Cell Biochem. 2021 Mar;476(3):1529-1540. doi: 10.1007/s11010-020-04002-3. Epub 2021 Jan 4. Mol Cell Biochem. 2021. PMID: 33394272 Review.
Cited by
-
Human Pannexin 1 Channel is NOT Phosphorylated by Src Tyrosine Kinase at Tyr199 and Tyr309.bioRxiv [Preprint]. 2024 Mar 18:2023.09.10.557063. doi: 10.1101/2023.09.10.557063. bioRxiv. 2024. Update in: Elife. 2024 May 23;13:RP95118. doi: 10.7554/eLife.95118. PMID: 38168229 Free PMC article. Updated. Preprint.
-
The role of maternal infections in neurodevelopmental psychiatric disorders: focus on the P2X7/NLRP3/IL-1β signalling pathway.J Neuroinflammation. 2025 Jul 26;22(1):192. doi: 10.1186/s12974-025-03509-0. J Neuroinflammation. 2025. PMID: 40713568 Free PMC article. Review.
-
Mechanosensitive Pannexin 1 Activity Is Modulated by Stomatin in Human Red Blood Cells.Int J Mol Sci. 2022 Aug 20;23(16):9401. doi: 10.3390/ijms23169401. Int J Mol Sci. 2022. PMID: 36012667 Free PMC article.
-
Mechanosensitive release of ATP in the urinary bladder mucosa.Purinergic Signal. 2025 Jun;21(3):413-428. doi: 10.1007/s11302-024-10063-6. Epub 2024 Nov 14. Purinergic Signal. 2025. PMID: 39541058 Free PMC article. Review.
-
New Panx-1 Blockers: Synthesis, Biological Evaluation and Molecular Dynamic Studies.Int J Mol Sci. 2022 Apr 27;23(9):4827. doi: 10.3390/ijms23094827. Int J Mol Sci. 2022. PMID: 35563213 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
