

The Unicellular Red Alga Cyanidioschyzon merolae-The Simplest Model of a Photosynthetic Eukaryote
- PMID: 33836072
- PMCID: PMC8504449
- DOI: 10.1093/pcp/pcab052
The Unicellular Red Alga Cyanidioschyzon merolae-The Simplest Model of a Photosynthetic Eukaryote
Abstract
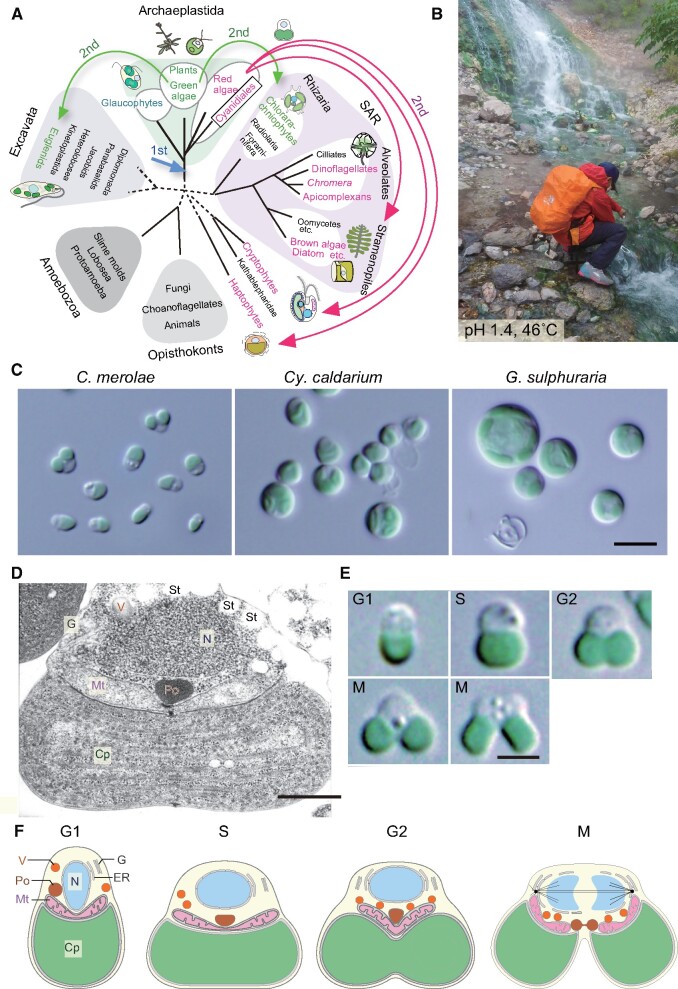
Several species of unicellular eukaryotic algae exhibit relatively simple genomic and cellular architecture. Laboratory cultures of these algae grow faster than plants and often provide homogeneous cellular populations exposed to an almost equal environment. These characteristics are ideal for conducting experiments at the cellular and subcellular levels. Many microalgal lineages have recently become genetically tractable, which have started to evoke new streams of studies. Among such algae, the unicellular red alga Cyanidioschyzon merolae is the simplest organism; it possesses the minimum number of membranous organelles, only 4,775 protein-coding genes in the nucleus, and its cell cycle progression can be highly synchronized with the diel cycle. These properties facilitate diverse omics analyses of cellular proliferation and structural analyses of the intracellular relationship among organelles. C. merolae cells lack a rigid cell wall and are thus relatively easily disrupted, facilitating biochemical analyses. Multiple chromosomal loci can be edited by highly efficient homologous recombination. The procedures for the inducible/repressive expression of a transgene or an endogenous gene in the nucleus and for chloroplast genome modification have also been developed. Here, we summarize the features and experimental techniques of C. merolae and provide examples of studies using this alga. From these studies, it is clear that C. merolae-either alone or in comparative and combinatory studies with other photosynthetic organisms-can provide significant insights into the biology of photosynthetic eukaryotes.
Keywords: Cyanidioschyzon merolae; Cyanidiales; Photosynthetic eukaryotes; Red algae.
© The Author(s) 2021. Published by Oxford University Press on behalf of Japanese Society of Plant Physiologists.
Figures


References
-
- Albertano P., Ciniglia C., Pinto G., Pollio A. (2000) The taxonomic position of Cyanidium, Cyanidioschyzon and Galdieria: an update. Hydrobiologia 433: 137–143.
-
- Antoshvili M., Caspy I., Hippler M., Nelson N. (2019) Structure and function of photosystem I in Cyanidioschyzon merolae. Photosynth. Res. 139: 499–508. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
