

Suppression of liquid-liquid phase separation by 1,6-hexanediol partially compromises the 3D genome organization in living cells
- PMID: 33836078
- PMCID: PMC8501969
- DOI: 10.1093/nar/gkab249
Suppression of liquid-liquid phase separation by 1,6-hexanediol partially compromises the 3D genome organization in living cells
Abstract
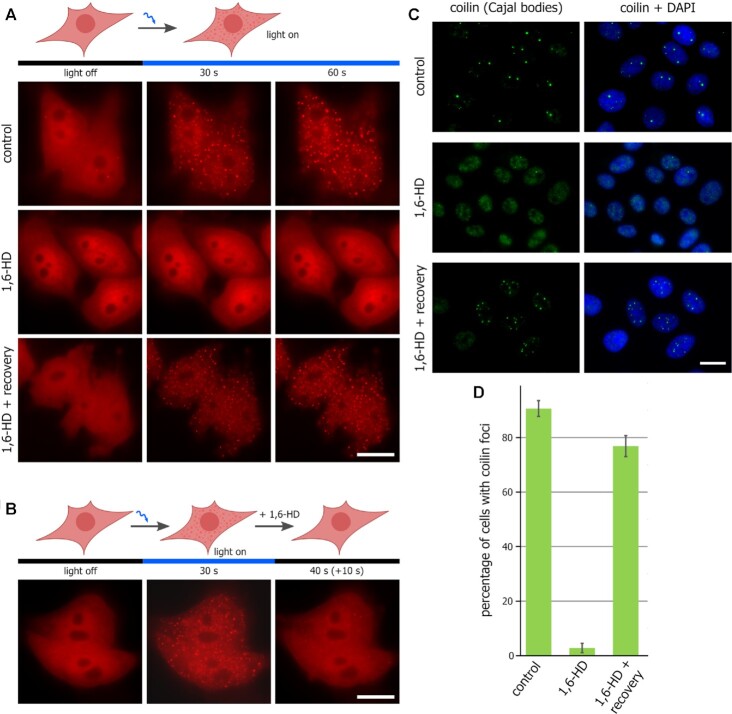
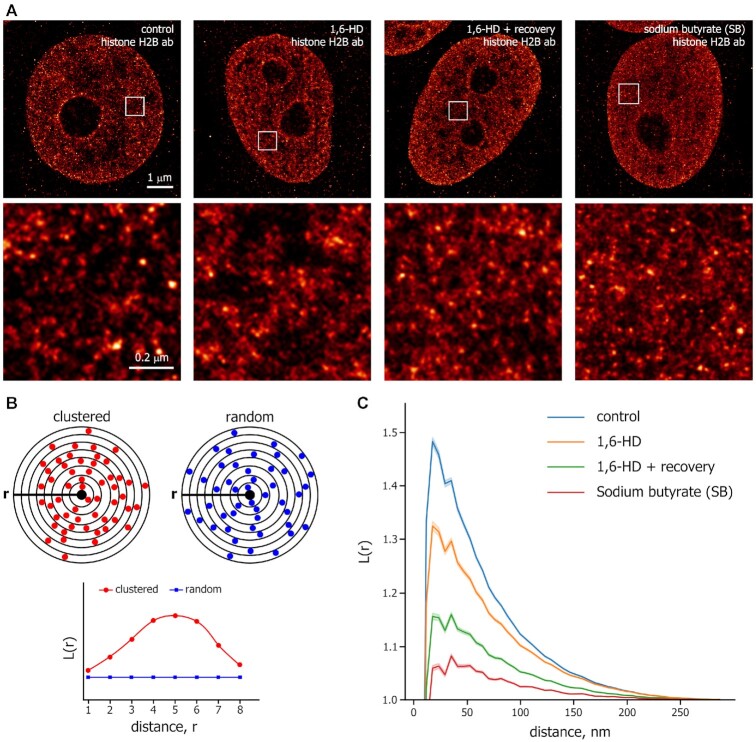
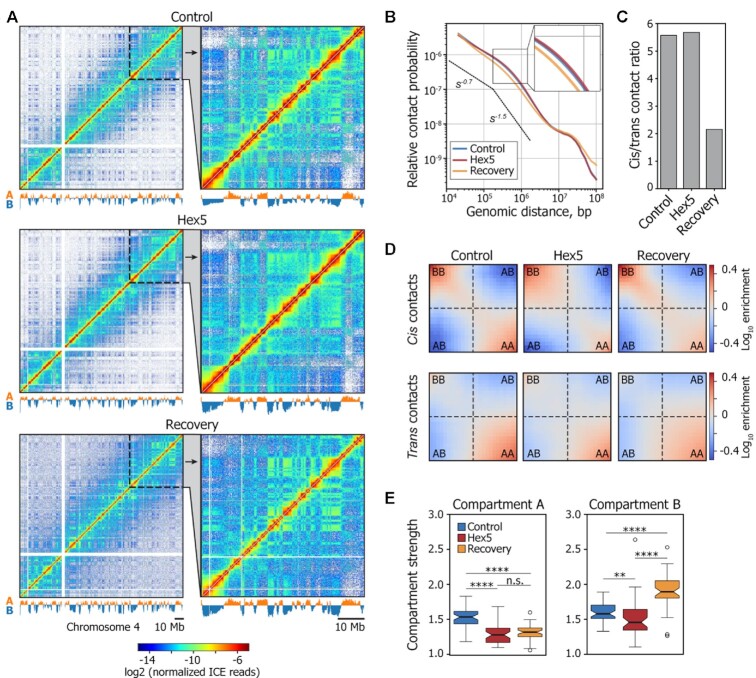
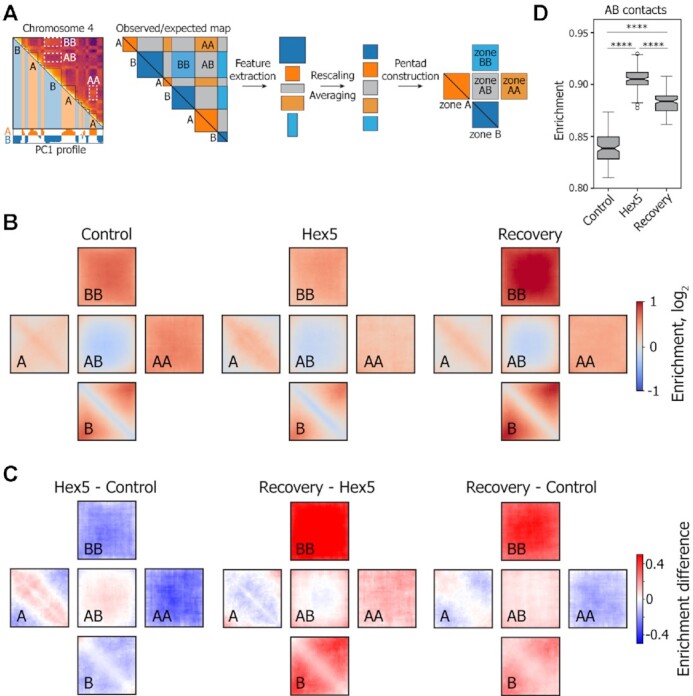
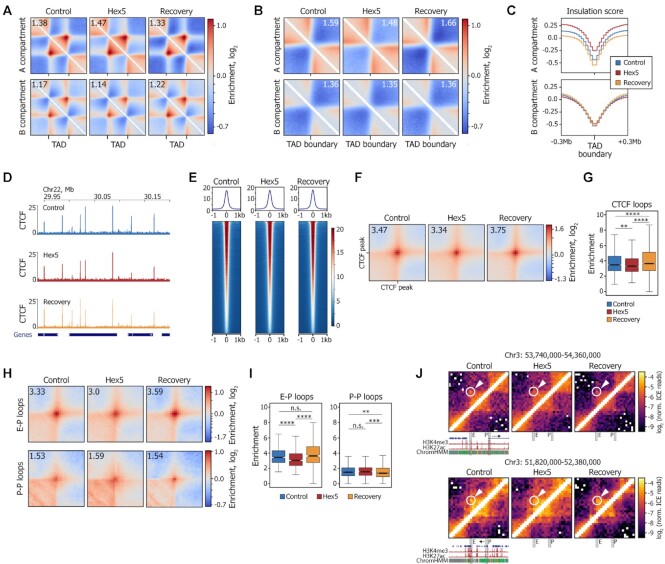
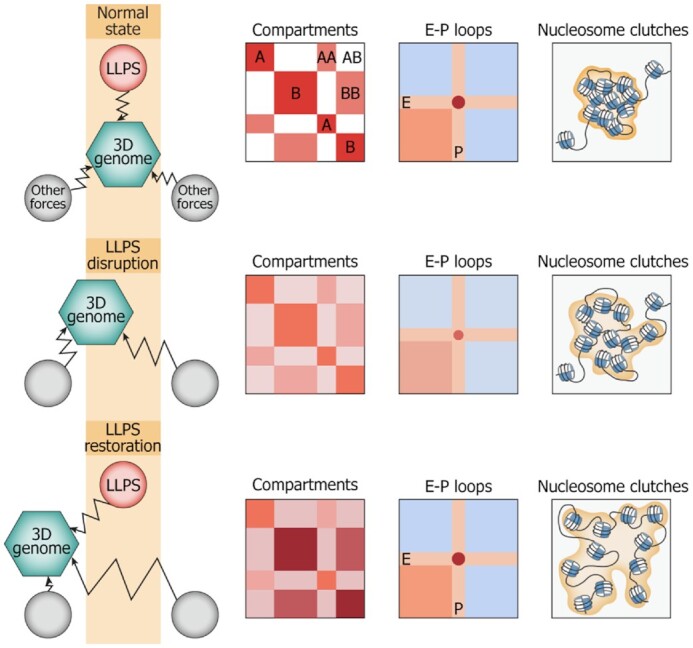
Liquid-liquid phase separation (LLPS) contributes to the spatial and functional segregation of molecular processes within the cell nucleus. However, the role played by LLPS in chromatin folding in living cells remains unclear. Here, using stochastic optical reconstruction microscopy (STORM) and Hi-C techniques, we studied the effects of 1,6-hexanediol (1,6-HD)-mediated LLPS disruption/modulation on higher-order chromatin organization in living cells. We found that 1,6-HD treatment caused the enlargement of nucleosome clutches and their more uniform distribution in the nuclear space. At a megabase-scale, chromatin underwent moderate but irreversible perturbations that resulted in the partial mixing of A and B compartments. The removal of 1,6-HD from the culture medium did not allow chromatin to acquire initial configurations, and resulted in more compact repressed chromatin than in untreated cells. 1,6-HD treatment also weakened enhancer-promoter interactions and TAD insulation but did not considerably affect CTCF-dependent loops. Our results suggest that 1,6-HD-sensitive LLPS plays a limited role in chromatin spatial organization by constraining its folding patterns and facilitating compartmentalization at different levels.
© The Author(s) 2021. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Similar articles
-
Time-dependent effect of 1,6-hexanediol on biomolecular condensates and 3D chromatin organization.Genome Biol. 2021 Aug 17;22(1):230. doi: 10.1186/s13059-021-02455-3. Genome Biol. 2021. PMID: 34404453 Free PMC article.
-
Quantifying the phase separation property of chromatin-associated proteins under physiological conditions using an anti-1,6-hexanediol index.Genome Biol. 2021 Aug 17;22(1):229. doi: 10.1186/s13059-021-02456-2. Genome Biol. 2021. PMID: 34404448 Free PMC article.
-
1,6-hexanediol rapidly immobilizes and condenses chromatin in living human cells.Life Sci Alliance. 2021 Feb 3;4(4):e202001005. doi: 10.26508/lsa.202001005. Print 2021 Apr. Life Sci Alliance. 2021. PMID: 33536240 Free PMC article.
-
Implications of liquid-liquid phase separation in plant chromatin organization and transcriptional control.Curr Opin Genet Dev. 2019 Apr;55:59-65. doi: 10.1016/j.gde.2019.06.003. Epub 2019 Jul 12. Curr Opin Genet Dev. 2019. PMID: 31306885 Review.
-
Nuclear compartments, genome folding, and enhancer-promoter communication.Int Rev Cell Mol Biol. 2015;315:183-244. doi: 10.1016/bs.ircmb.2014.11.004. Epub 2015 Feb 9. Int Rev Cell Mol Biol. 2015. PMID: 25708464 Review.
Cited by
-
Pseudogenes and Liquid Phase Separation in Epigenetic Expression.Front Oncol. 2022 Jul 8;12:912282. doi: 10.3389/fonc.2022.912282. eCollection 2022. Front Oncol. 2022. PMID: 35875144 Free PMC article. Review.
-
Liquid condensates: a new barrier to loop extrusion?Cell Mol Life Sci. 2025 Feb 20;82(1):80. doi: 10.1007/s00018-024-05559-8. Cell Mol Life Sci. 2025. PMID: 39976773 Free PMC article. Review.
-
DNA methylation insulates genic regions from CTCF loops near nuclear speckles.Elife. 2025 Sep 3;13:RP102930. doi: 10.7554/eLife.102930. Elife. 2025. PMID: 40900123 Free PMC article.
-
Phase Separation in Chromatin Organization and Human Diseases.Int J Mol Sci. 2025 May 28;26(11):5156. doi: 10.3390/ijms26115156. Int J Mol Sci. 2025. PMID: 40507965 Free PMC article. Review.
-
Phase Separation in the Nucleus and at the Nuclear Periphery during Post-Mitotic Nuclear Envelope Reformation.Cells. 2022 May 25;11(11):1749. doi: 10.3390/cells11111749. Cells. 2022. PMID: 35681444 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
