

Discovery of fungal oligosaccharide-oxidising flavo-enzymes with previously unknown substrates, redox-activity profiles and interplay with LPMOs
- PMID: 33837197
- PMCID: PMC8035211
- DOI: 10.1038/s41467-021-22372-0
Discovery of fungal oligosaccharide-oxidising flavo-enzymes with previously unknown substrates, redox-activity profiles and interplay with LPMOs
Abstract
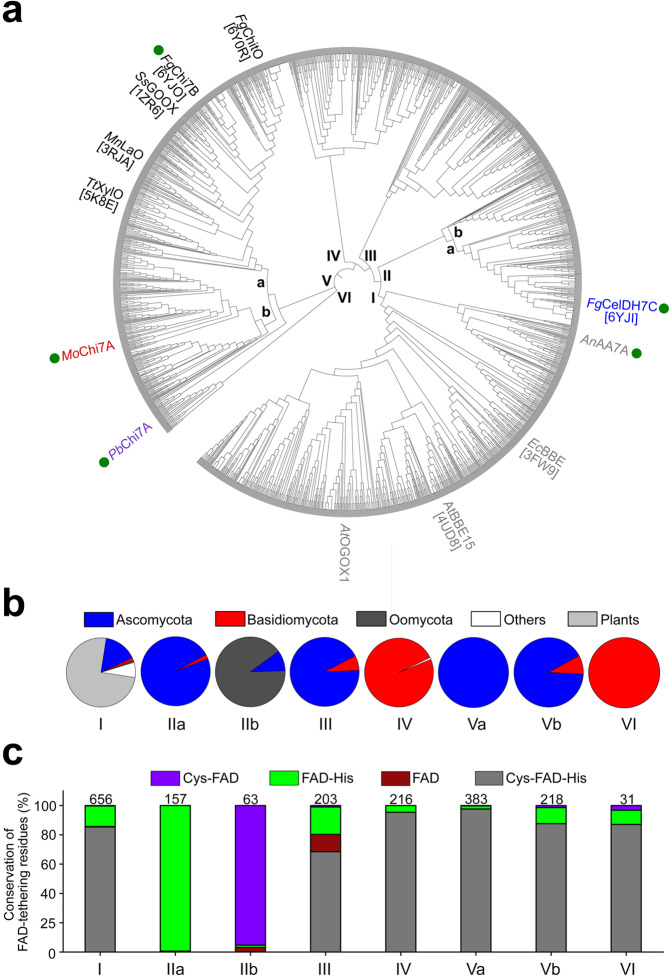
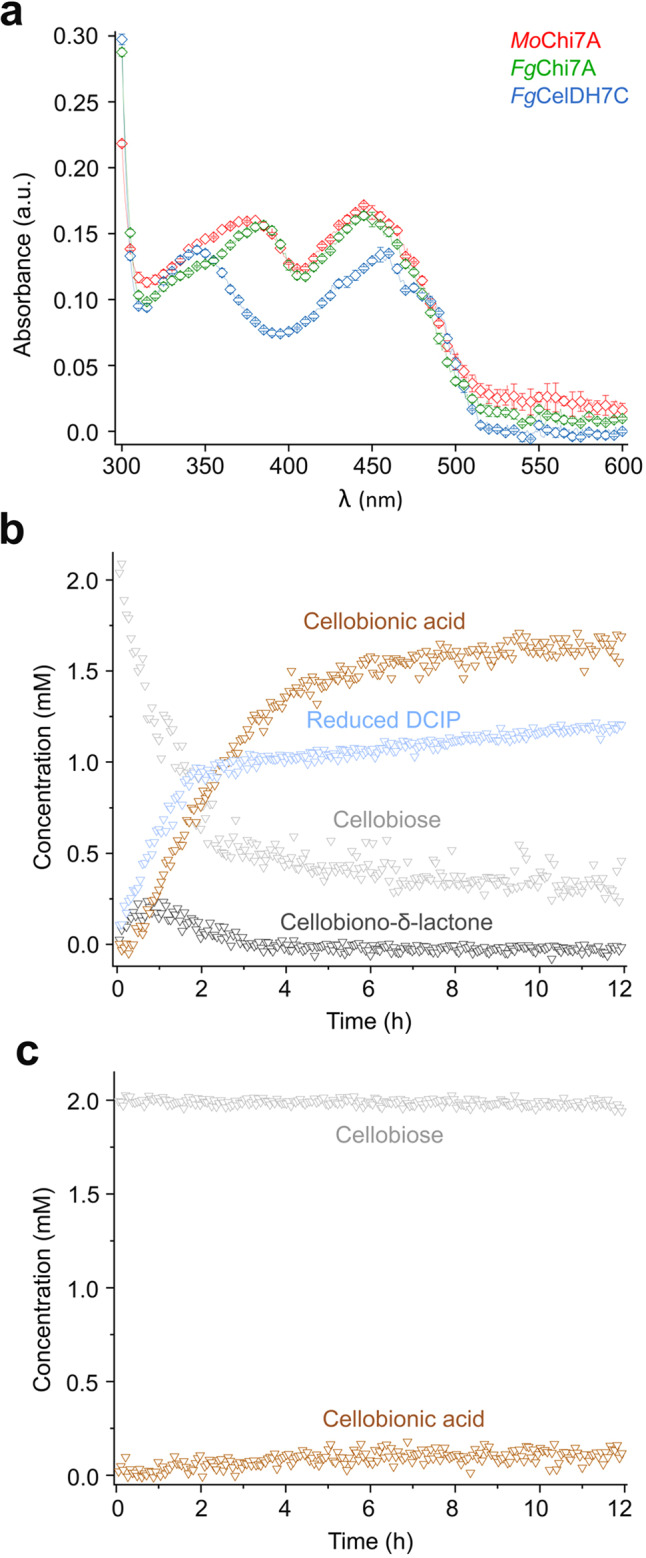
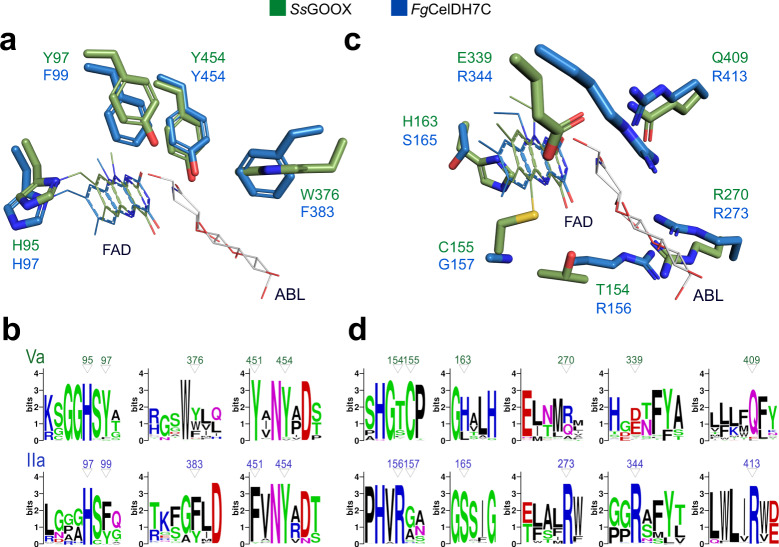
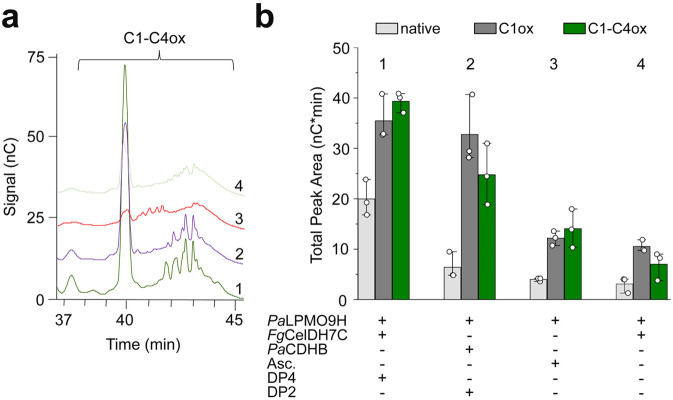
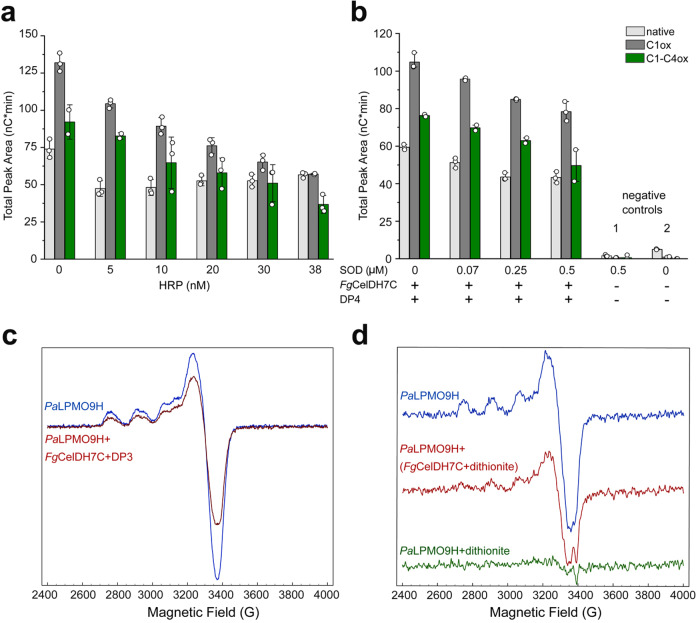
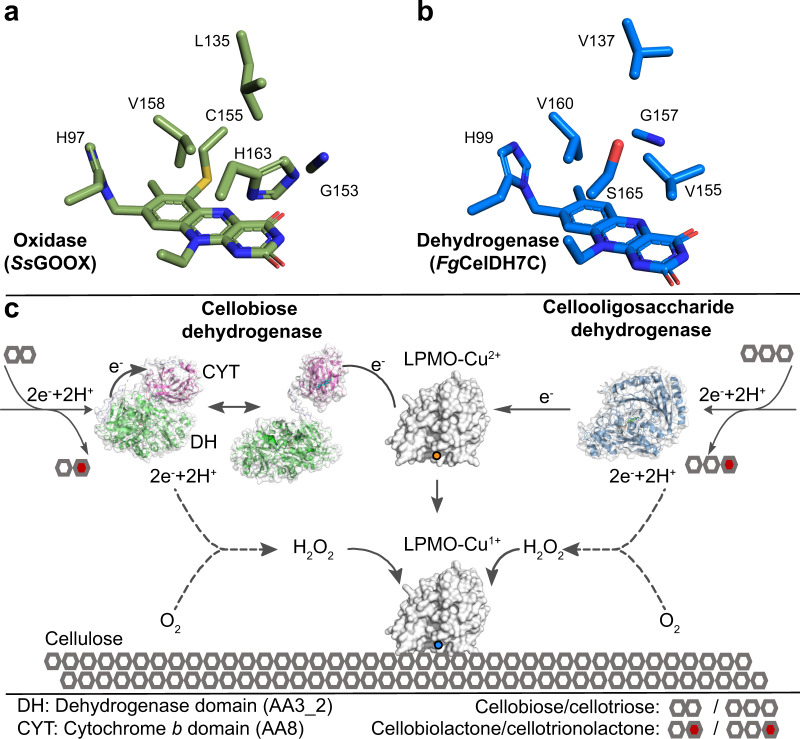
Oxidative plant cell-wall processing enzymes are of great importance in biology and biotechnology. Yet, our insight into the functional interplay amongst such oxidative enzymes remains limited. Here, a phylogenetic analysis of the auxiliary activity 7 family (AA7), currently harbouring oligosaccharide flavo-oxidases, reveals a striking abundance of AA7-genes in phytopathogenic fungi and Oomycetes. Expression of five fungal enzymes, including three from unexplored clades, expands the AA7-substrate range and unveils a cellooligosaccharide dehydrogenase activity, previously unknown within AA7. Sequence and structural analyses identify unique signatures distinguishing the strict dehydrogenase clade from canonical AA7 oxidases. The discovered dehydrogenase directly is able to transfer electrons to an AA9 lytic polysaccharide monooxygenase (LPMO) and fuel cellulose degradation by LPMOs without exogenous reductants. The expansion of redox-profiles and substrate range highlights the functional diversity within AA7 and sets the stage for harnessing AA7 dehydrogenases to fine-tune LPMO activity in biotechnological conversion of plant feedstocks.
Conflict of interest statement
A provisional patent has been filed based on data from this paper by M.H.M., J.G.B. and M.A.H. The other authors declare that they have no competing interests.
Figures
Similar articles
-
Functional characterization of cellulose-degrading AA9 lytic polysaccharide monooxygenases and their potential exploitation.Appl Microbiol Biotechnol. 2020 Apr;104(8):3229-3243. doi: 10.1007/s00253-020-10467-5. Epub 2020 Feb 19. Appl Microbiol Biotechnol. 2020. PMID: 32076777 Review.
-
Lytic polysaccharide monooxygenases and other oxidative enzymes are abundantly secreted by Aspergillus nidulans grown on different starches.Biotechnol Biofuels. 2016 Sep 1;9(1):187. doi: 10.1186/s13068-016-0604-0. eCollection 2016. Biotechnol Biofuels. 2016. PMID: 27588040 Free PMC article.
-
The discovery of novel LPMO families with a new Hidden Markov model.BMC Res Notes. 2017 Feb 21;10(1):105. doi: 10.1186/s13104-017-2429-8. BMC Res Notes. 2017. PMID: 28222763 Free PMC article.
-
A Lytic Polysaccharide Monooxygenase with Broad Xyloglucan Specificity from the Brown-Rot Fungus Gloeophyllum trabeum and Its Action on Cellulose-Xyloglucan Complexes.Appl Environ Microbiol. 2016 Oct 27;82(22):6557-6572. doi: 10.1128/AEM.01768-16. Print 2016 Nov 15. Appl Environ Microbiol. 2016. PMID: 27590806 Free PMC article.
-
Lytic Polysaccharide Monooxygenases: The Microbial Power Tool for Lignocellulose Degradation.Trends Plant Sci. 2016 Nov;21(11):926-936. doi: 10.1016/j.tplants.2016.07.012. Epub 2016 Aug 12. Trends Plant Sci. 2016. PMID: 27527668 Review.
Cited by
-
Deletion of AA9 Lytic Polysaccharide Monooxygenases Impacts A. nidulans Secretome and Growth on Lignocellulose.Microbiol Spectr. 2022 Jun 29;10(3):e0212521. doi: 10.1128/spectrum.02125-21. Epub 2022 Jun 6. Microbiol Spectr. 2022. PMID: 35658600 Free PMC article.
-
Investigating lytic polysaccharide monooxygenase-assisted wood cell wall degradation with microsensors.Nat Commun. 2022 Oct 21;13(1):6258. doi: 10.1038/s41467-022-33963-w. Nat Commun. 2022. PMID: 36271009 Free PMC article.
-
Expanding the catalytic landscape of metalloenzymes with lytic polysaccharide monooxygenases.Nat Rev Chem. 2024 Feb;8(2):106-119. doi: 10.1038/s41570-023-00565-z. Epub 2024 Jan 10. Nat Rev Chem. 2024. PMID: 38200220 Review.
-
Challenges of Biomass Utilization for Bioenergy in a Climate Change Scenario.Biology (Basel). 2021 Dec 6;10(12):1277. doi: 10.3390/biology10121277. Biology (Basel). 2021. PMID: 34943192 Free PMC article. Review.
-
Genome and secretome insights: unravelling the lignocellulolytic potential of Myceliophthora verrucosa for enhanced hydrolysis of lignocellulosic biomass.Arch Microbiol. 2024 Apr 27;206(5):236. doi: 10.1007/s00203-024-03974-w. Arch Microbiol. 2024. PMID: 38676717
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
