

Spatial and temporal scales of dopamine transmission
- PMID: 33837376
- PMCID: PMC8220193
- DOI: 10.1038/s41583-021-00455-7
Spatial and temporal scales of dopamine transmission
Abstract
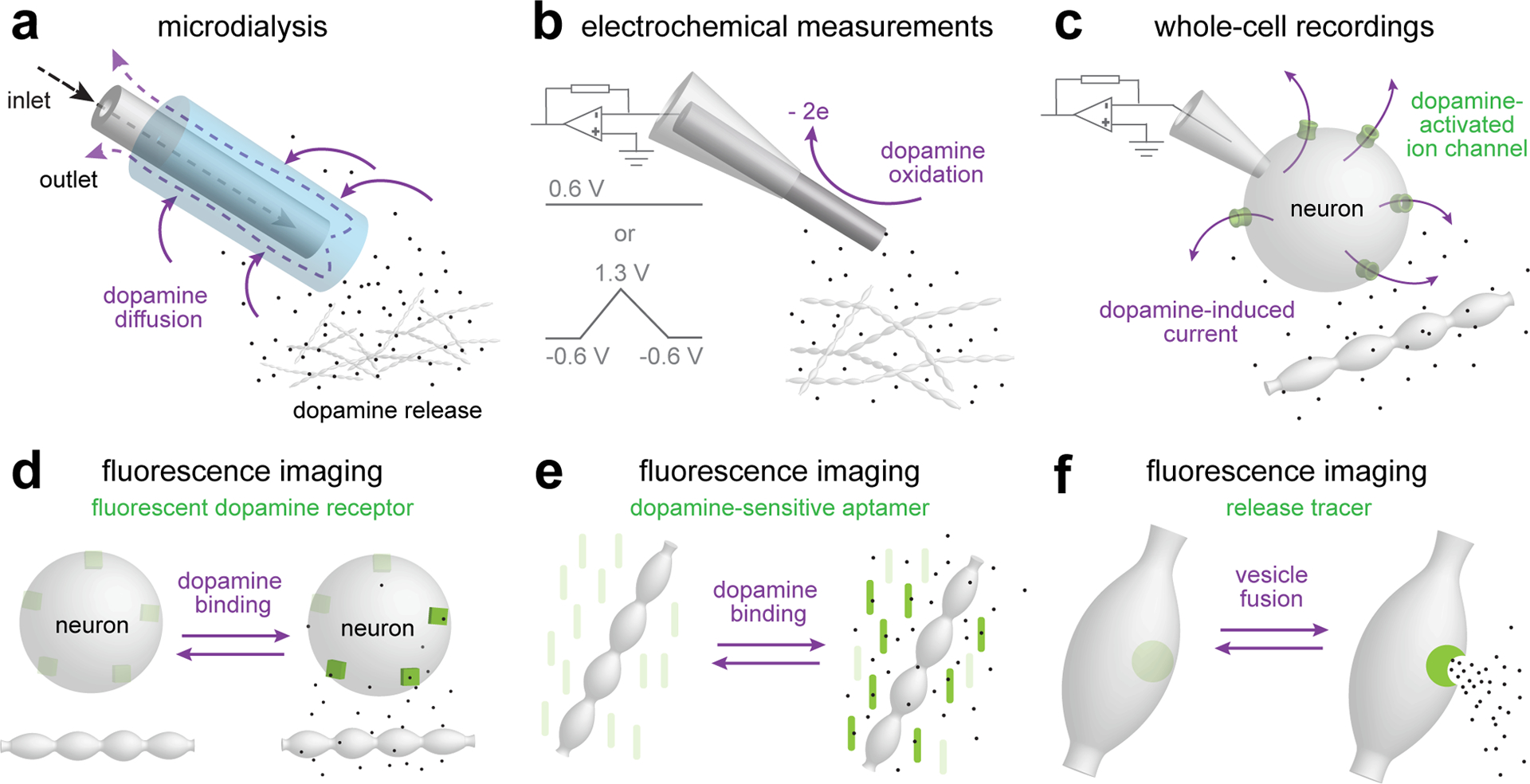
Dopamine is a prototypical neuromodulator that controls circuit function through G protein-coupled receptor signalling. Neuromodulators are volume transmitters, with release followed by diffusion for widespread receptor activation on many target cells. Yet, we are only beginning to understand the specific organization of dopamine transmission in space and time. Although some roles of dopamine are mediated by slow and diffuse signalling, recent studies suggest that certain dopamine functions necessitate spatiotemporal precision. Here, we review the literature describing dopamine signalling in the striatum, including its release mechanisms and receptor organization. We then propose the domain-overlap model, in which release and receptors are arranged relative to one another in micrometre-scale structures. This architecture is different from both point-to-point synaptic transmission and the widespread organization that is often proposed for neuromodulation. It enables the activation of receptor subsets that are within micrometre-scale domains of release sites during baseline activity and broader receptor activation with domain overlap when firing is synchronized across dopamine neuron populations. This signalling structure, together with the properties of dopamine release, may explain how switches in firing modes support broad and dynamic roles for dopamine and may lead to distinct pathway modulation.
Figures





References
-
- Grillner S & Robertson B The Basal Ganglia Over 500 Million Years. Curr. Biol 26, R1088–R1100 (2016). - PubMed
-
- Sawin ER, Ranganathan R & Horvitz HRC elegans Locomotory Rate Is Modulated by the Environment through a Dopaminergic Pathway and by Experience through a Serotonergic Pathway. Neuron 26, 619–631 (2000). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
