

The multidomain architecture of a bacteriophage endolysin enables intramolecular synergism and regulation of bacterial lysis
- PMID: 33838182
- PMCID: PMC8144678
- DOI: 10.1016/j.jbc.2021.100639
The multidomain architecture of a bacteriophage endolysin enables intramolecular synergism and regulation of bacterial lysis
Abstract
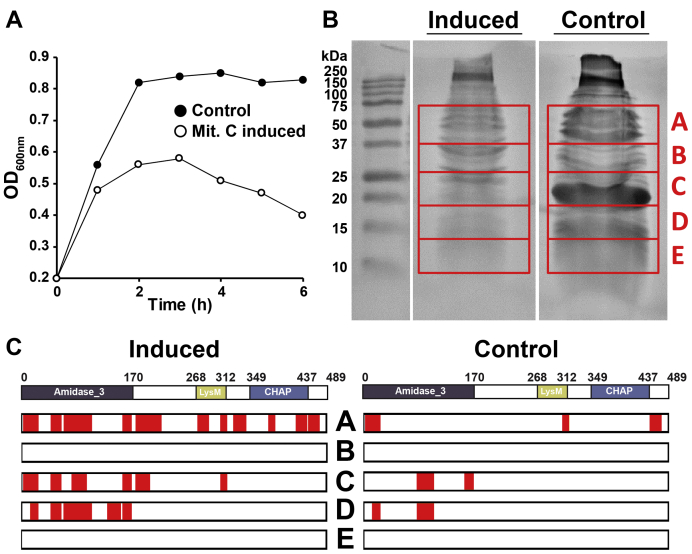
Endolysins are peptidoglycan hydrolases produced at the end of the bacteriophage (phage) replication cycle to lyse the host cell. Endolysins in Gram-positive phages come in a variety of multimodular forms that combine different catalytic and cell wall binding domains. However, the reason why phages adopt endolysins with such complex multidomain architecture is not well understood. In this study, we used the Streptococcus dysgalactiae phage endolysin PlySK1249 as a model to investigate the role of multidomain architecture in phage-induced bacterial lysis and lysis regulation. PlySK1249 consists of an amidase (Ami) domain that lyses bacterial cells, a nonbacteriolytic endopeptidase (CHAP) domain that acts as a dechaining enzyme, and a central LysM cell wall binding domain. We observed that the Ami and CHAP domains synergized for peptidoglycan digestion and bacteriolysis in the native enzyme or when expressed individually and reunified. The CHAP endopeptidase resolved complex polymers of stem-peptides to dimers and helped the Ami domain to digest peptidoglycan to completion. We also found that PlySK1249 was subject to proteolytic cleavage by host cell wall proteases both in vitro and after phage induction. Cleavage disconnected the different domains by hydrolyzing their linker regions, thus hindering their bacteriolytic cooperation and possibly modulating the lytic activity of the enzyme. PlySK1249 cleavage by cell-wall-associated proteases may represent another example of phage adaptation toward the use of existing bacterial regulation mechanism for their own advantage. In addition, understanding more thoroughly the multidomain interplay of PlySK1249 broadens our knowledge on the ideal architecture of therapeutic antibacterial endolysins.
Keywords: PlySK1249; bacteriophage; endolysin; intramolecular synergism; lysis regulation; proteolysis.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures






References
-
- Nelson D.C., Schmelcher M., Rodriguez-Rubio L., Klumpp J., Pritchard D.G., Dong S., Donovan D.M. Endolysins as antimicrobials. Adv. Virus Res. 2012;83:299–365. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
