

Interferon regulatory factor 8 regulates expression of acid ceramidase and infection susceptibility in cystic fibrosis
- PMID: 33839155
- PMCID: PMC8113888
- DOI: 10.1016/j.jbc.2021.100650
Interferon regulatory factor 8 regulates expression of acid ceramidase and infection susceptibility in cystic fibrosis
Abstract
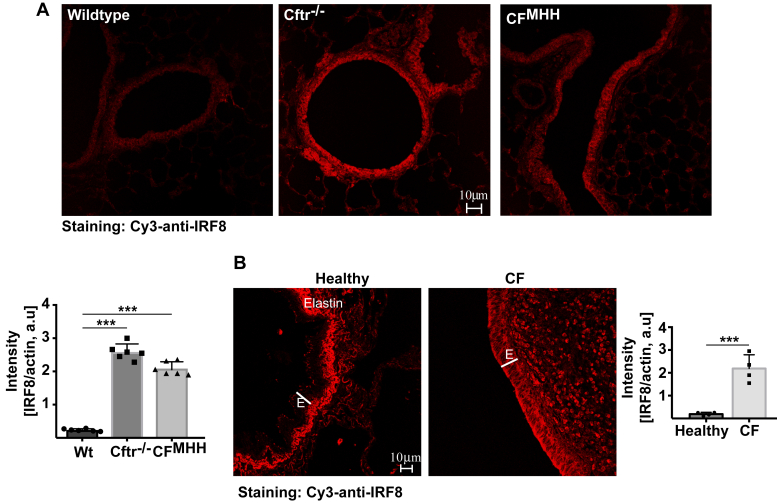
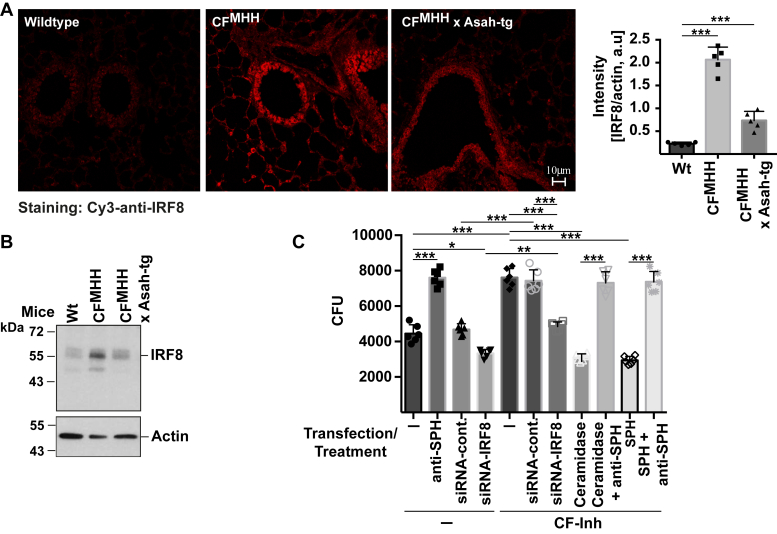
Most patients with cystic fibrosis (CF) suffer from acute and chronic pulmonary infections with bacterial pathogens, which often determine their life quality and expectancy. Previous studies have demonstrated a downregulation of the acid ceramidase in CF epithelial cells resulting in an increase of ceramide and a decrease of sphingosine. Sphingosine kills many bacterial pathogens, and the downregulation of sphingosine seems to determine the infection susceptibility of cystic fibrosis mice and patients. It is presently unknown how deficiency of the cystic fibrosis transmembrane conductance regulator (CFTR) connects to a marked downregulation of the acid ceramidase in human and murine CF epithelial cells. Here, we employed quantitative PCR, western blot analysis, and enzyme activity measurements to study the role of IRF8 for acid ceramidase regulation. We report that genetic deficiency or functional inhibition of CFTR/Cftr results in an upregulation of interferon regulatory factor 8 (IRF8) and a concomitant downregulation of acid ceramidase expression with CF and an increase of ceramide and a reduction of sphingosine levels in tracheal and bronchial epithelial cells from both human individuals or mice. CRISPR/Cas9- or siRNA-mediated downregulation of IRF8 prevented changes of acid ceramidase, ceramide, and sphingosine in CF epithelial cells and restored resistance to Pseudomonas aeruginosa infections, which is one of the most important and common pathogens in lung infection of patients with CF. These studies indicate that CFTR deficiency causes a downregulation of acid ceramidase via upregulation of IRF8, which is a central pathway to control infection susceptibility of CF cells.
Keywords: Pseudomonas aeruginosa; acid ceramidase; ceramide; cystic fibrosis; interferon response factor-8; sphingosine.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interests The authors declare no competing financial interests. M. B., not related to this work: investigator-led research grants from Pfizer and Roche Diagnostics; speaker fees paid to Newcastle University from Novartis, Roche Diagnostics, and TEVA. Travel expenses to educational meetings: Boehringer Ingelheim and Vertex Pharmaceuticals.
Figures





References
-
- Riordan J.R., Rommens J.M., Kerem B., Alon N., Rozmahel R., Grzelczak Z., Zielenski J., Lok S., Plavsic N., Chou J.L., Drumm M.L., Iannuzzi M.C., Collins F.S., Tsui L.C. Identification of the cystic fibrosis gene: Cloning and characterization of complementary DNA. Science. 1989;245:1066–1073. - PubMed
-
- Kerem B., Rommens J.M., Buchanan J.A., Markiewicz D., Cox T.K., Chakravarti A., Buchwald M., Tsui L.C. Identification of the cystic fibrosis gene: Genetic analysis. Science. 1989;245:1073–1080. - PubMed
-
- Elborn J.S. Cystic fibrosis. Lancet. 2016;388:2519–2531. - PubMed
-
- CF Foundation . Cystic Fibrosis Foundation; Bethesda, MD: 2019. Patient Registry Annual Report.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
