

The interplay between DNA and histone methylation: molecular mechanisms and disease implications
- PMID: 33844406
- PMCID: PMC8097341
- DOI: 10.15252/embr.202051803
The interplay between DNA and histone methylation: molecular mechanisms and disease implications
Abstract
Methylation of cytosine in CpG dinucleotides and histone lysine and arginine residues is a chromatin modification that critically contributes to the regulation of genome integrity, replication, and accessibility. A strong correlation exists between the genome-wide distribution of DNA and histone methylation, suggesting an intimate relationship between these epigenetic marks. Indeed, accumulating literature reveals complex mechanisms underlying the molecular crosstalk between DNA and histone methylation. These in vitro and in vivo discoveries are further supported by the finding that genes encoding DNA- and histone-modifying enzymes are often mutated in overlapping human diseases. Here, we summarize recent advances in understanding how DNA and histone methylation cooperate to maintain the cellular epigenomic landscape. We will also discuss the potential implication of these insights for understanding the etiology of, and developing biomarkers and therapies for, human congenital disorders and cancers that are driven by chromatin abnormalities.
Keywords: DNA methylation; cancer; chromatin; developmental disorder; histone methylation.
© 2021 The Authors.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
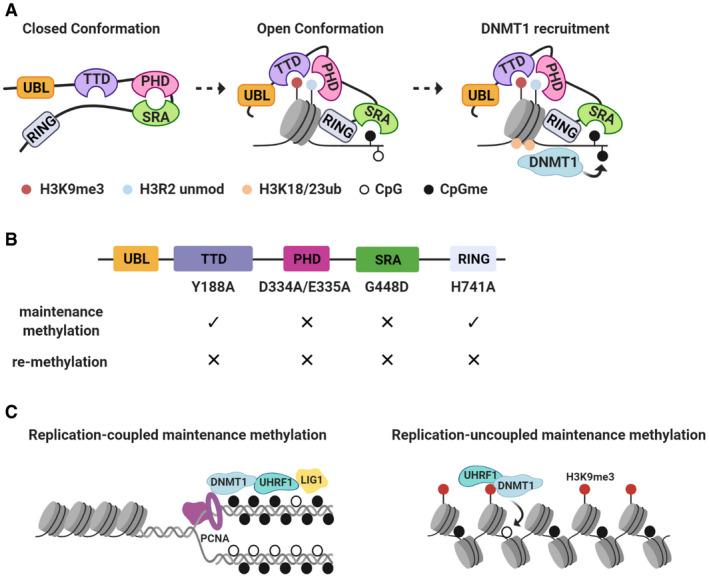
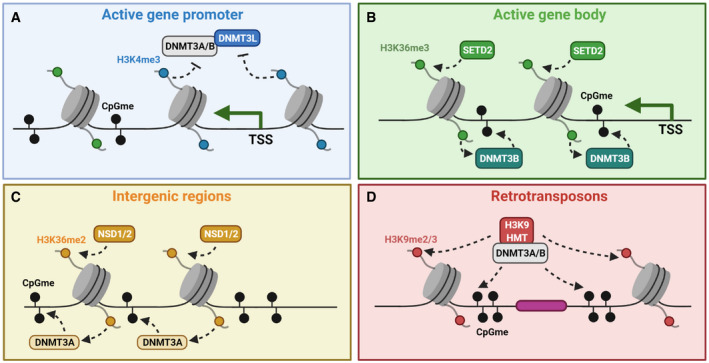
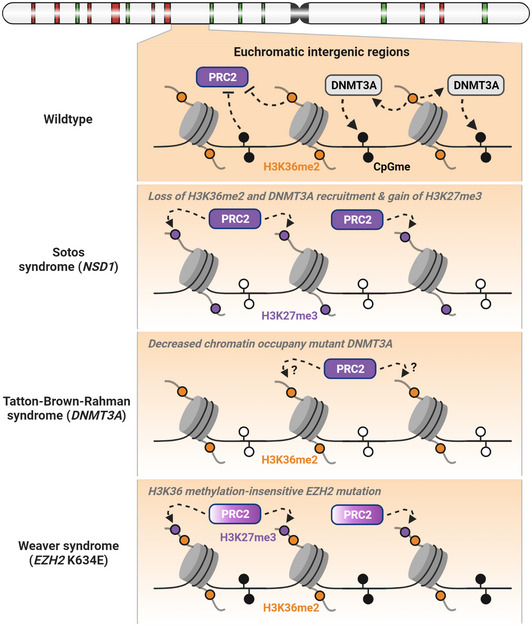
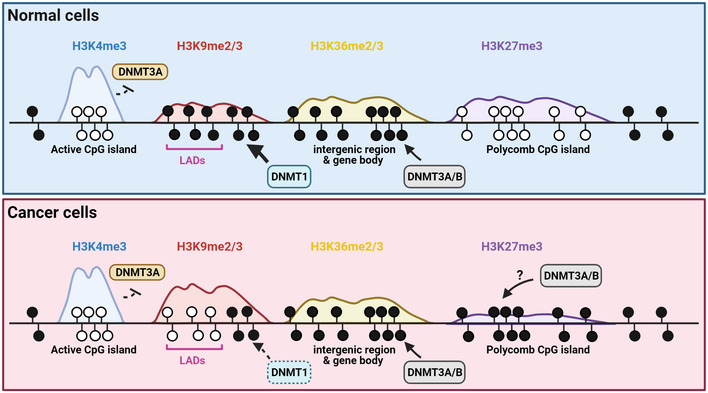
References
-
- Ai S, Xiong H, Li CC, Luo Y, Shi Q, Liu Y, Yu X, Li C, He A (2019) Profiling chromatin states using single‐cell itChIP‐seq. Nat Cell Biol 21: 1164–1172 - PubMed
-
- Aref‐Eshghi E, Bourque DK, Kerkhof J, Carere DA, Ainsworth P, Sadikovic B, Armour CM, Lin H (2019) Genome‐wide DNA methylation and RNA analyses enable reclassification of two variants of uncertain significance in a patient with clinical Kabuki syndrome. Hum Mutat 40: 1684–1689 - PubMed
-
- Aref‐Eshghi E, Rodenhiser DI, Schenkel LC, Lin H, Skinner C, Ainsworth P, Paré G, Hood RL, Bulman DE, Kernohan KD et al (2018) Genomic DNA methylation signatures enable concurrent diagnosis and clinical genetic variant classification in neurodevelopmental syndromes. Am J Hum Genet 102: 156–174 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
