

Reovirus directly engages integrin to recruit clathrin for entry into host cells
- PMID: 33846319
- PMCID: PMC8041799
- DOI: 10.1038/s41467-021-22380-0
Reovirus directly engages integrin to recruit clathrin for entry into host cells
Erratum in
-
Author Correction: Reovirus directly engages integrin to recruit clathrin for entry into host cells.Nat Commun. 2021 May 13;12(1):2946. doi: 10.1038/s41467-021-23537-7. Nat Commun. 2021. PMID: 33986270 Free PMC article. No abstract available.
Abstract
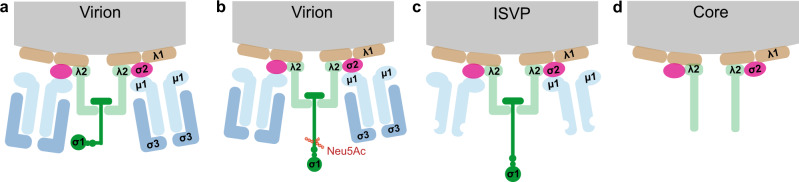
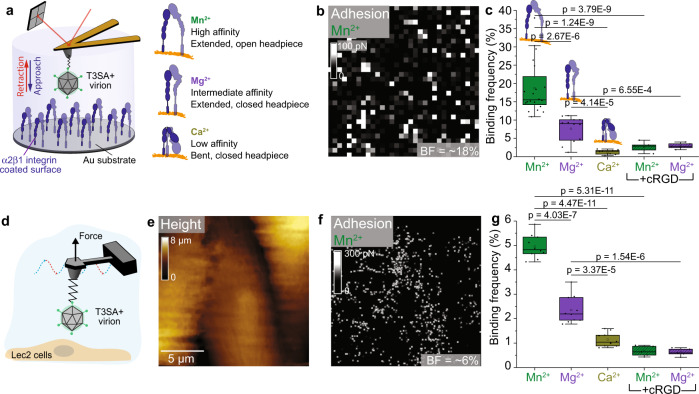
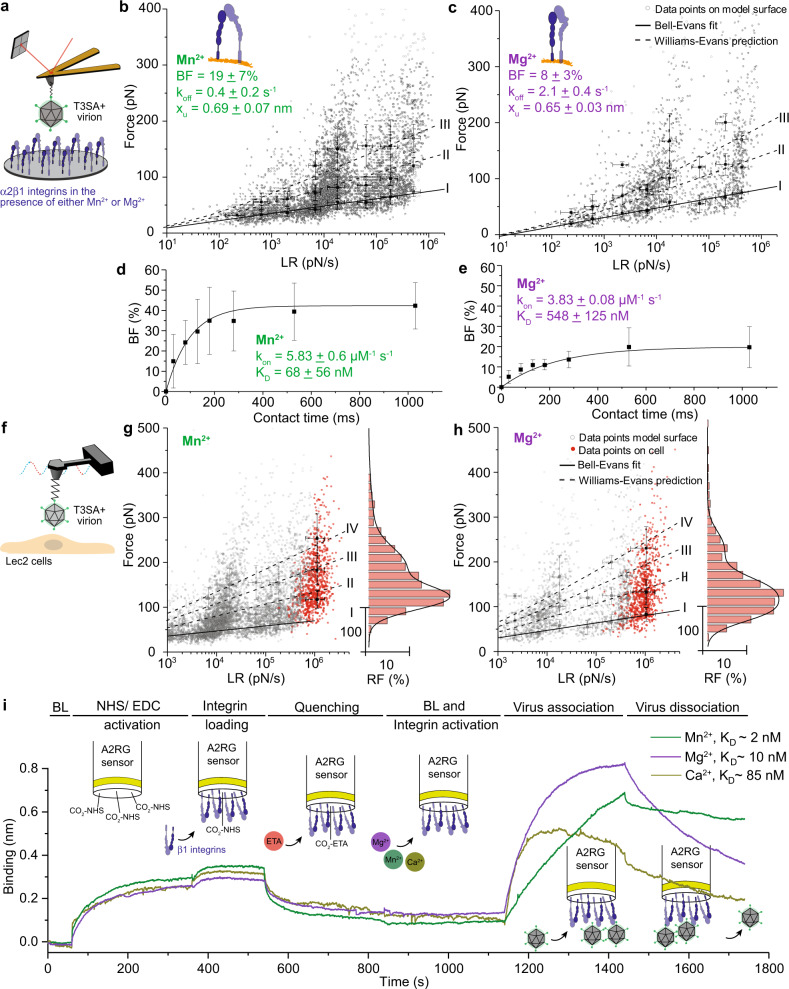
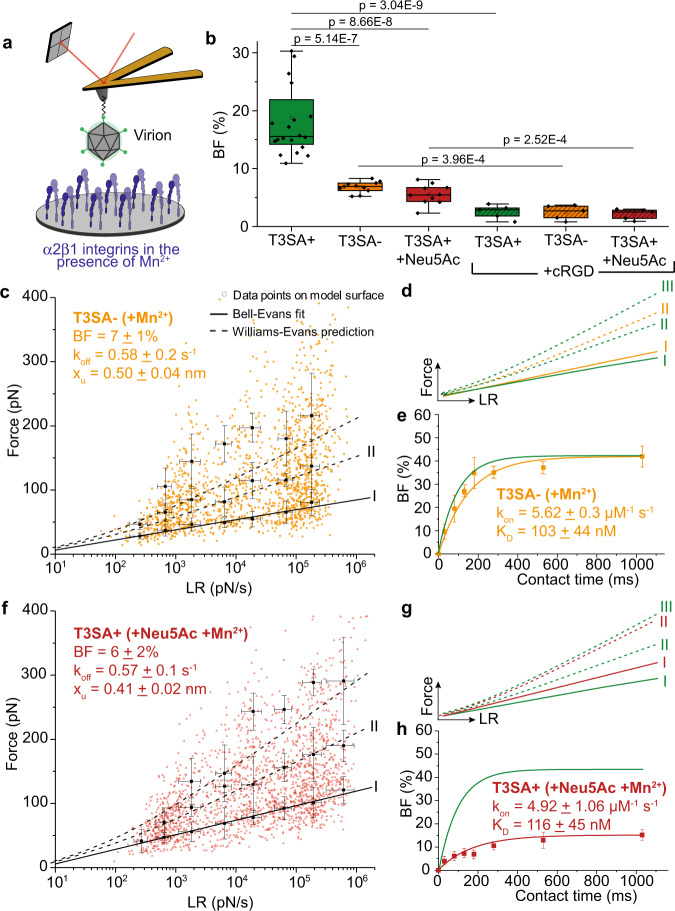
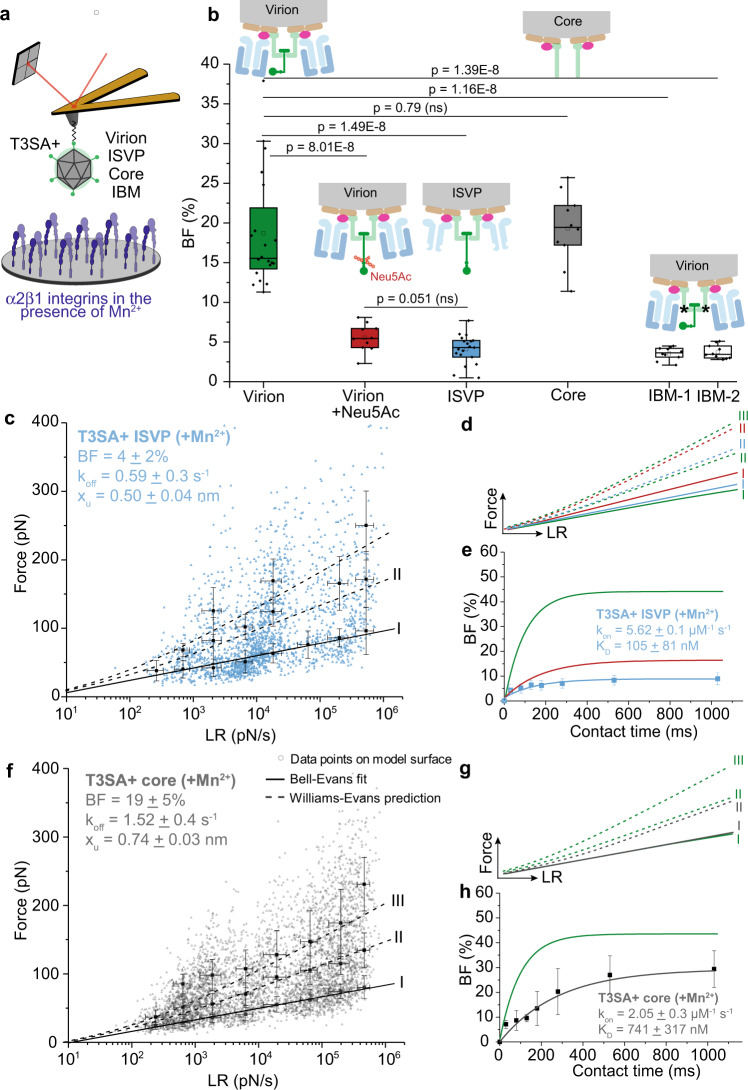
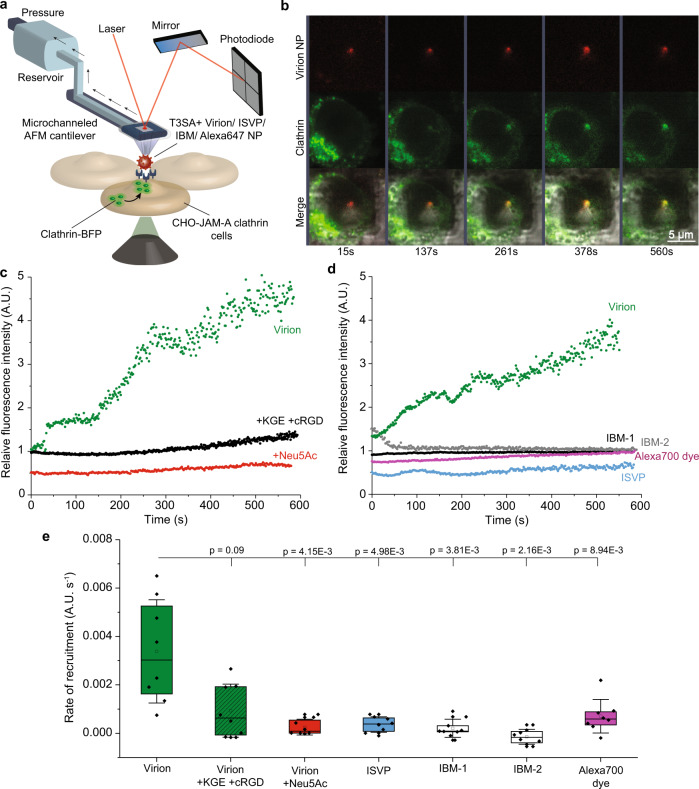
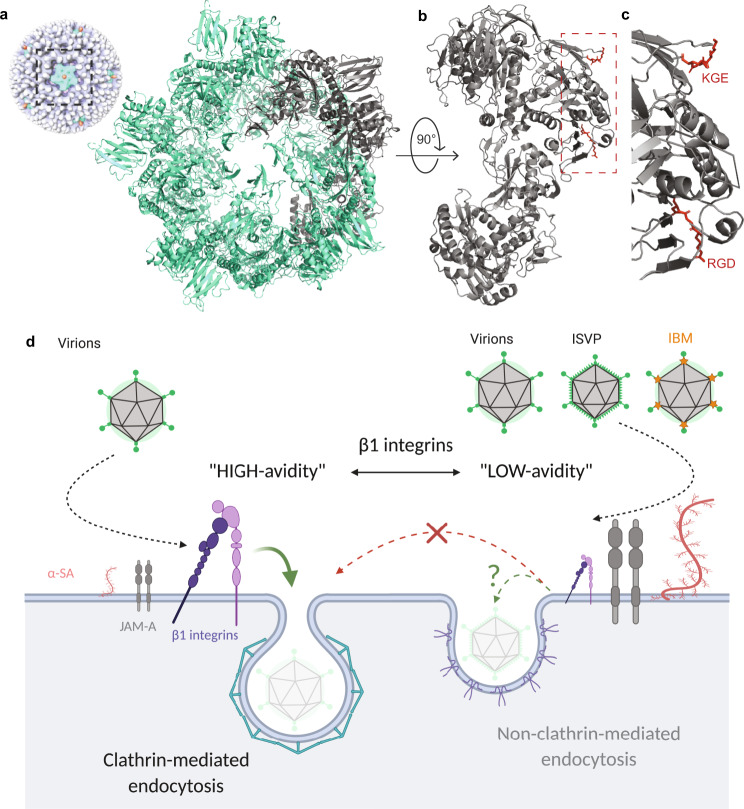
Reovirus infection requires the concerted action of viral and host factors to promote cell entry. After interaction of reovirus attachment protein σ1 with cell-surface carbohydrates and proteinaceous receptors, additional host factors mediate virus internalization. In particular, β1 integrin is required for endocytosis of reovirus virions following junctional adhesion molecule A (JAM-A) binding. While integrin-binding motifs in the surface-exposed region of reovirus capsid protein λ2 are thought to mediate integrin interaction, evidence for direct β1 integrin-reovirus interactions and knowledge of how integrins function to mediate reovirus entry is lacking. Here, we use single-virus force spectroscopy and confocal microscopy to discover a direct interaction between reovirus and β1 integrins. Comparison of interactions between reovirus disassembly intermediates as well as mutants and β1 integrin show that λ2 is the integrin ligand. Finally, using fluidic force microscopy, we demonstrate a functional role for β1 integrin interaction in promoting clathrin recruitment to cell-bound reovirus. Our study demonstrates a direct interaction between reovirus and β1 integrins and offers insights into the mechanism of reovirus cell entry. These results provide new perspectives for the development of efficacious antiviral therapeutics and the engineering of improved viral gene delivery and oncolytic vectors.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
NPXY motifs in the beta1 integrin cytoplasmic tail are required for functional reovirus entry.J Virol. 2008 Apr;82(7):3181-91. doi: 10.1128/JVI.01612-07. Epub 2008 Jan 23. J Virol. 2008. PMID: 18216114 Free PMC article.
-
Beta1 integrin mediates internalization of mammalian reovirus.J Virol. 2006 Mar;80(6):2760-70. doi: 10.1128/JVI.80.6.2760-2770.2006. J Virol. 2006. PMID: 16501085 Free PMC article.
-
From touchdown to transcription: the reovirus cell entry pathway.Curr Top Microbiol Immunol. 2010;343:91-119. doi: 10.1007/82_2010_32. Curr Top Microbiol Immunol. 2010. PMID: 20397070 Free PMC article. Review.
-
Reovirus receptors, cell entry, and proapoptotic signaling.Adv Exp Med Biol. 2013;790:42-71. doi: 10.1007/978-1-4614-7651-1_3. Adv Exp Med Biol. 2013. PMID: 23884585 Free PMC article. Review.
-
Structural Insights into Reovirus σ1 Interactions with Two Neutralizing Antibodies.J Virol. 2017 Jan 31;91(4):e01621-16. doi: 10.1128/JVI.01621-16. Print 2017 Feb 15. J Virol. 2017. PMID: 27928010 Free PMC article.
Cited by
-
The reovirus variant RP116 is oncolytic in immunocompetent models and generates reduced neutralizing antibodies to Type 3 Dearing.Mol Ther Oncol. 2024 Jun 29;32(3):200846. doi: 10.1016/j.omton.2024.200846. eCollection 2024 Sep 19. Mol Ther Oncol. 2024. PMID: 39354956 Free PMC article.
-
Host Subcellular Organelles: Targets of Viral Manipulation.Int J Mol Sci. 2024 Jan 29;25(3):1638. doi: 10.3390/ijms25031638. Int J Mol Sci. 2024. PMID: 38338917 Free PMC article. Review.
-
Exosomal transmission of viruses, a two-edged biological sword.Cell Commun Signal. 2023 Jan 23;21(1):19. doi: 10.1186/s12964-022-01037-5. Cell Commun Signal. 2023. PMID: 36691072 Free PMC article. Review.
-
An overview of the role of Niemann-pick C1 (NPC1) in viral infections and inhibition of viral infections through NPC1 inhibitor.Cell Commun Signal. 2023 Dec 14;21(1):352. doi: 10.1186/s12964-023-01376-x. Cell Commun Signal. 2023. PMID: 38098077 Free PMC article. Review.
-
Viruses Binding to Host Receptors Interacts with Autophagy.Int J Mol Sci. 2023 Feb 8;24(4):3423. doi: 10.3390/ijms24043423. Int J Mol Sci. 2023. PMID: 36834833 Free PMC article. Review.
References
-
- Smith AE, Helenius A. How viruses enter animal cells. Science. 2004;304:237–242. - PubMed
-
- Dermody, T. & Tyler, K. Introduction to viruses and viral diseases. Mandell, Douglas, and Bennett’s principles and practice of infectious diseases, 6th edn. Churchill Livingstone, New York, NY, 1729–1742 (2005).
-
- Coffey MC, Strong JE, Forsyth PA, Lee PW. Reovirus therapy of tumors with activated Ras pathway. Science. 1998;282:1332–1334. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
