

Protein acylation by saturated very long chain fatty acids and endocytosis are involved in necroptosis
- PMID: 33848465
- PMCID: PMC8529612
- DOI: 10.1016/j.chembiol.2021.03.012
Protein acylation by saturated very long chain fatty acids and endocytosis are involved in necroptosis
Abstract
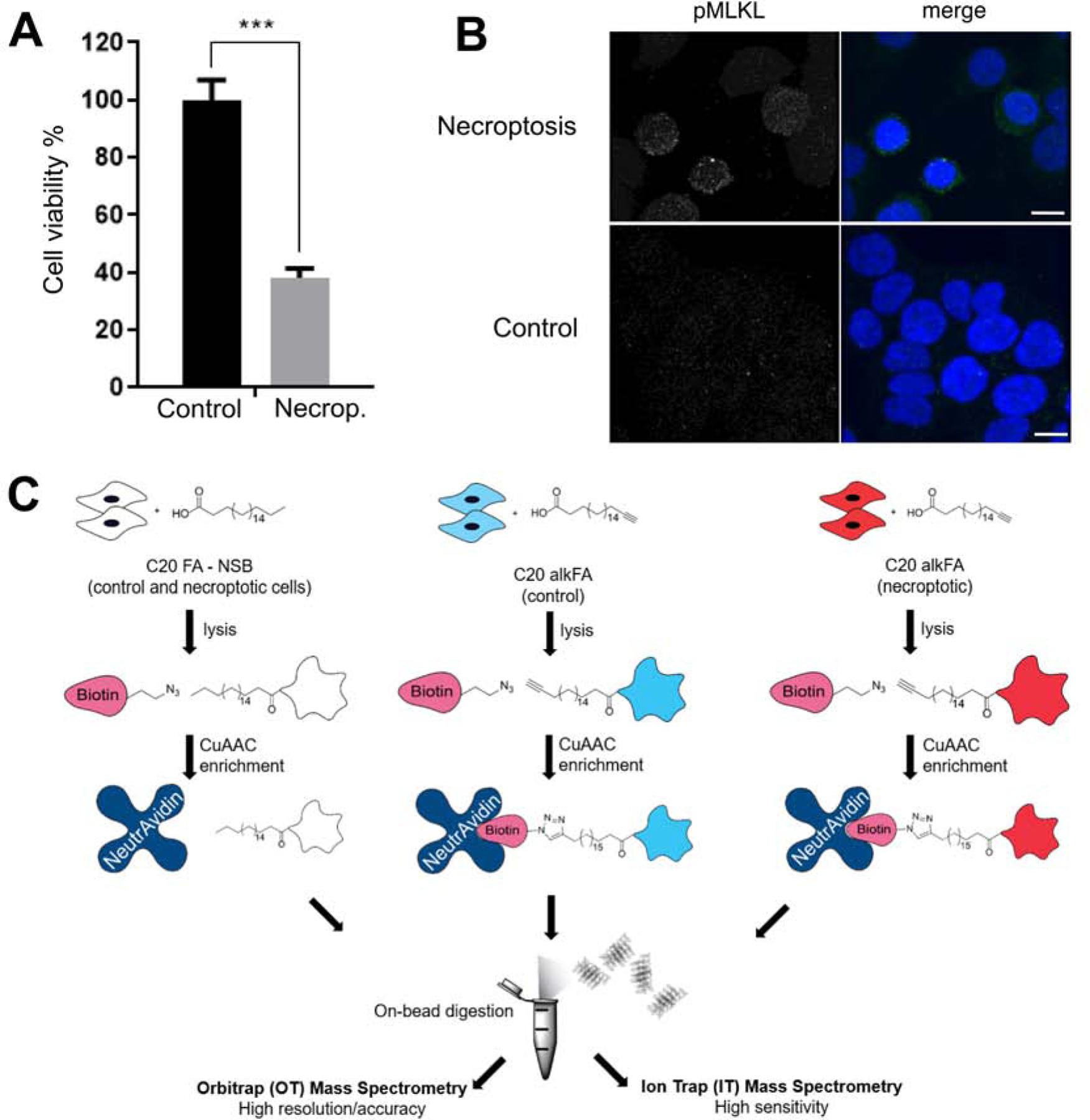
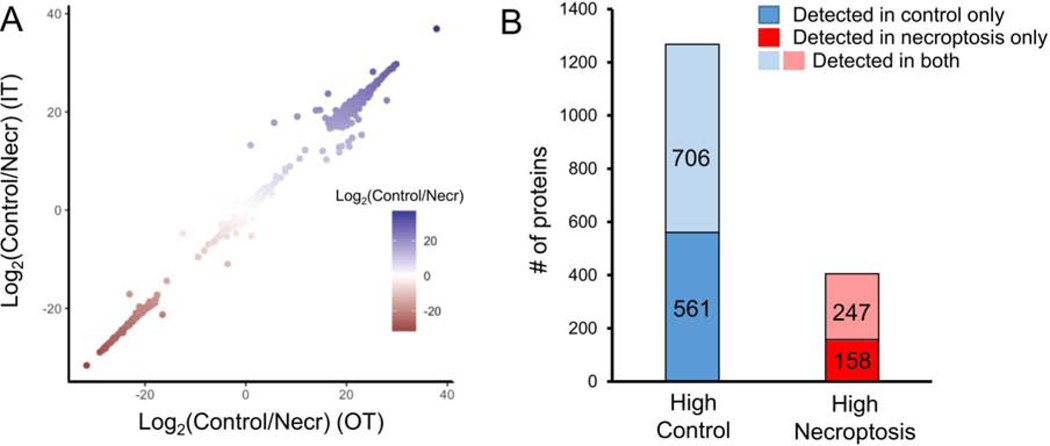
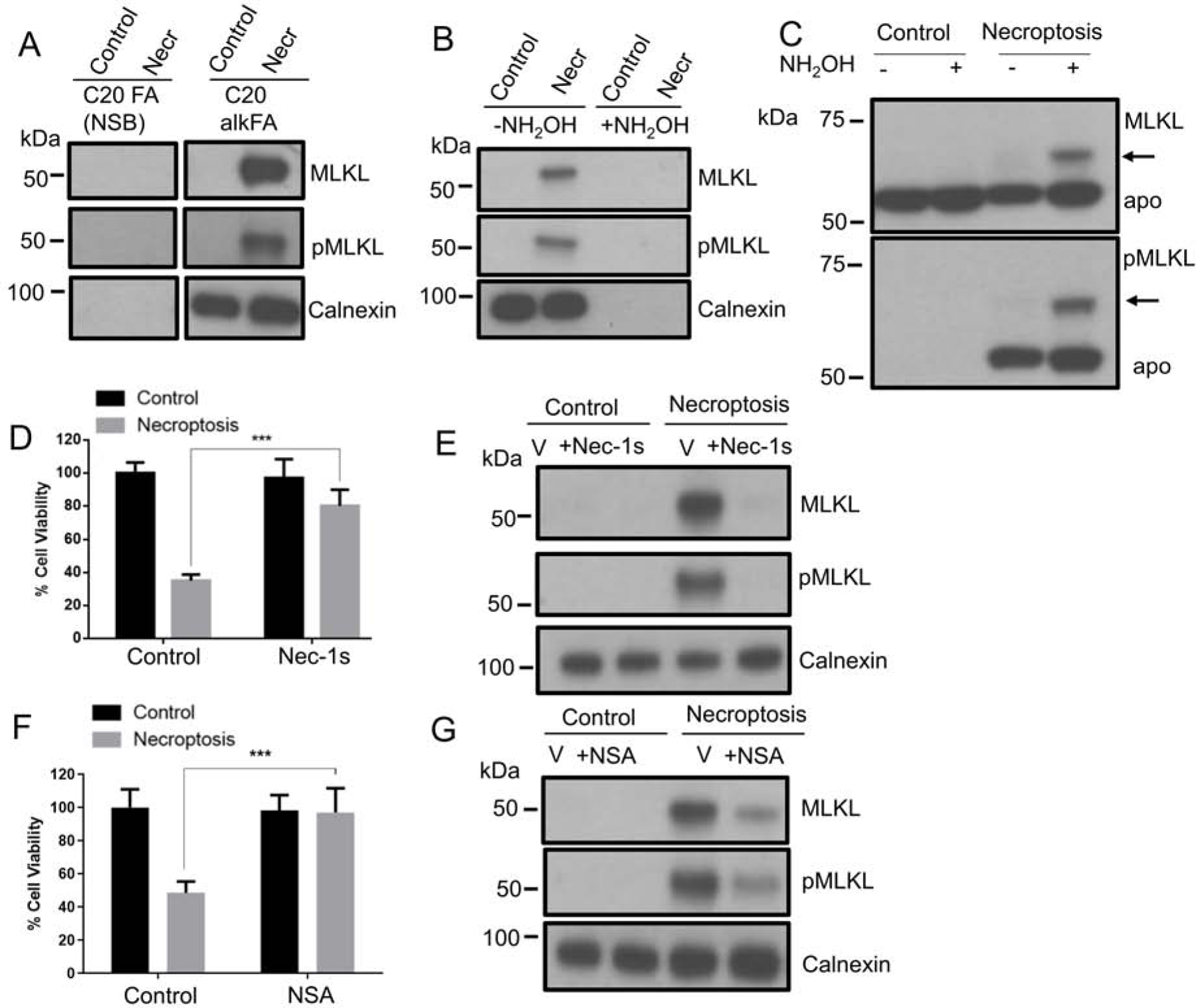
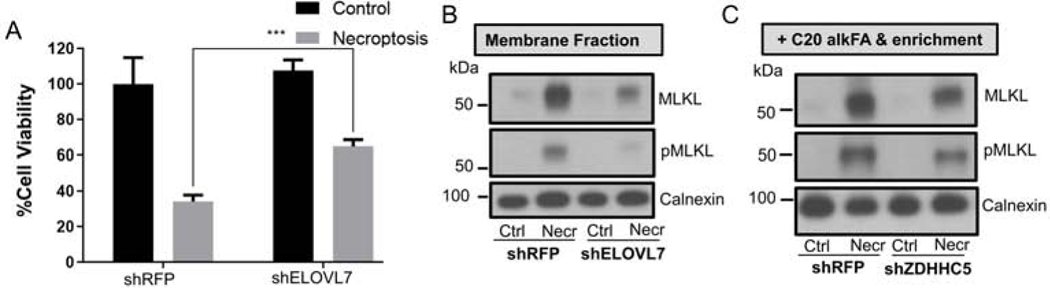
Necroptosis is a form of cell death characterized by receptor-interacting protein kinase activity and plasma membrane permeabilization via mixed-lineage kinase-like protein (MLKL). This permeabilization is responsible for the inflammatory properties of necroptosis. We previously showed that very long chain fatty acids (VLCFAs) are functionally involved in necroptosis, potentially through protein fatty acylation. Here, we define the scope of protein acylation by saturated VLCFAs during necroptosis. We show that MLKL and phosphoMLKL, key for membrane permeabilization, are exclusively acylated during necroptosis. Reducing the levels of VLCFAs decreases their membrane recruitment, suggesting that acylation by VLCFAs contributes to their membrane localization. Acylation of phosphoMLKL occurs downstream of phosphorylation and oligomerization and appears to be, in part, mediated by ZDHHC5 (a palmitoyl transferase). We also show that disruption of endosomal trafficking increases cell viability during necroptosis, possibly by preventing recruitment, or removal, of phosphoMLKL from the plasma membrane.
Keywords: VLCFA; ZDHHC5; endocytosis; lipids; necroptosis; programmed cell death; protein acylation; vesicular trafficking.
Copyright © 2021 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interest.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
