

Evolution of sexual development and sexual dimorphism in insects
- PMID: 33848958
- PMCID: PMC8364864
- DOI: 10.1016/j.gde.2021.02.011
Evolution of sexual development and sexual dimorphism in insects
Abstract
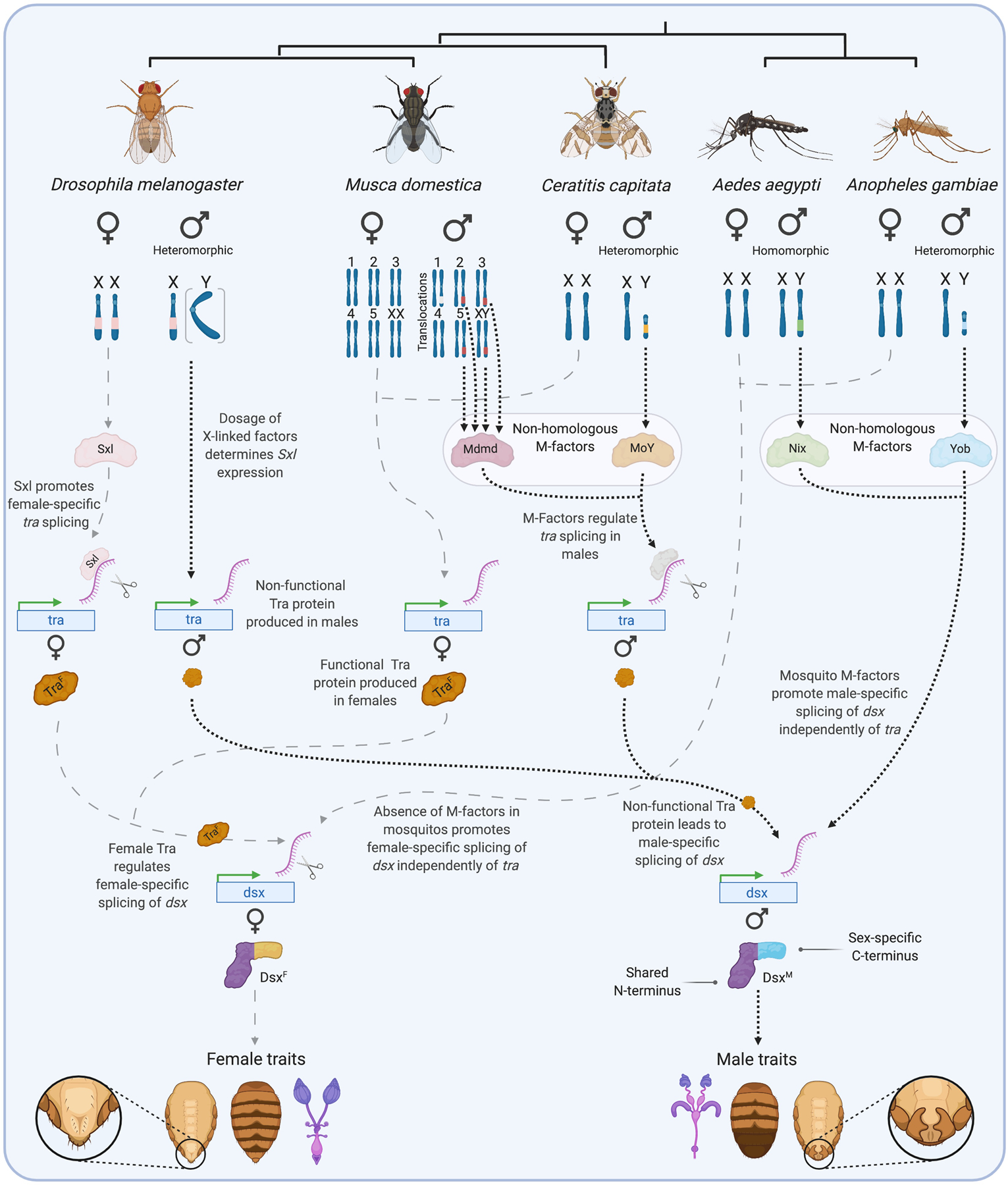
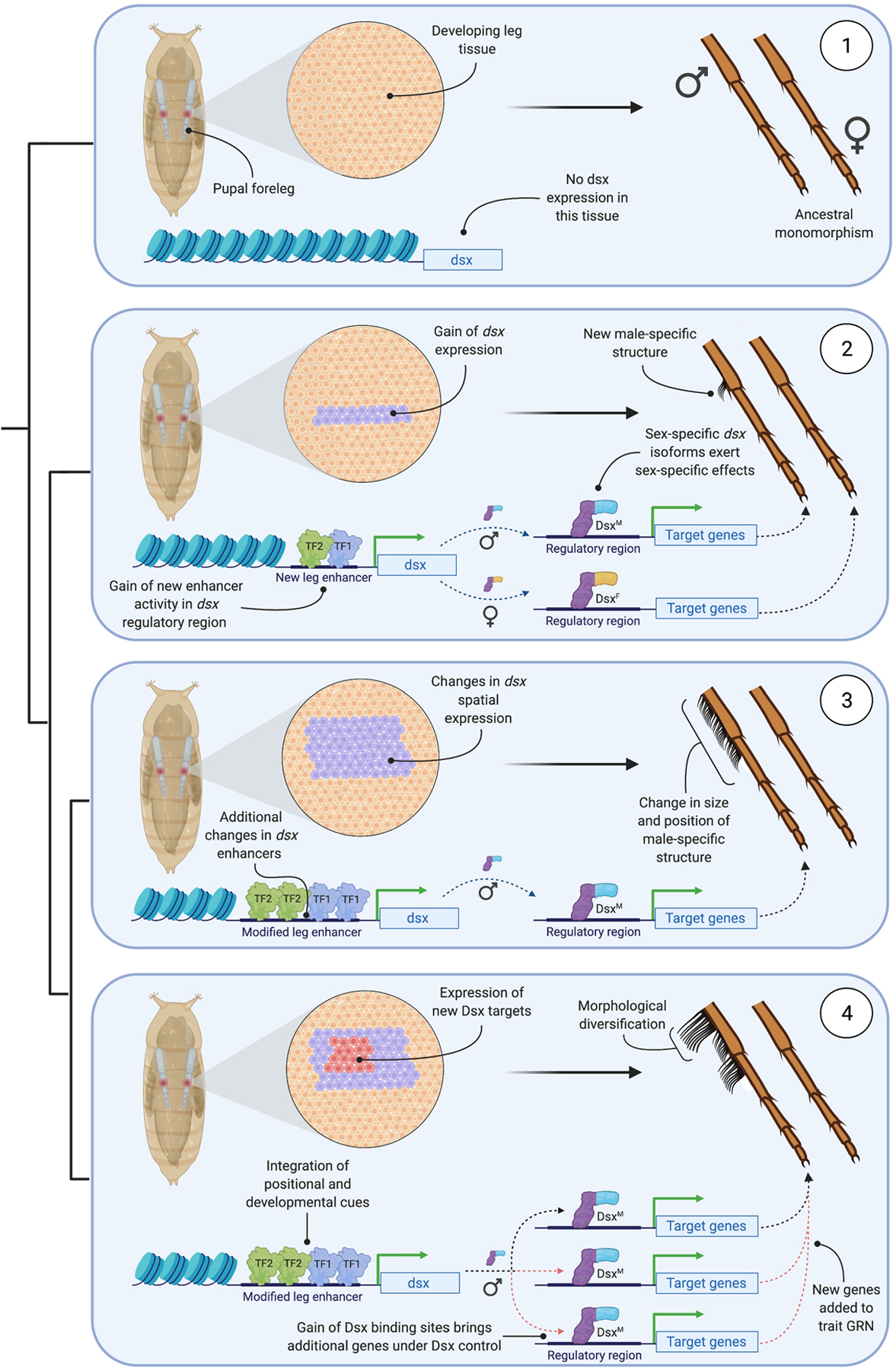
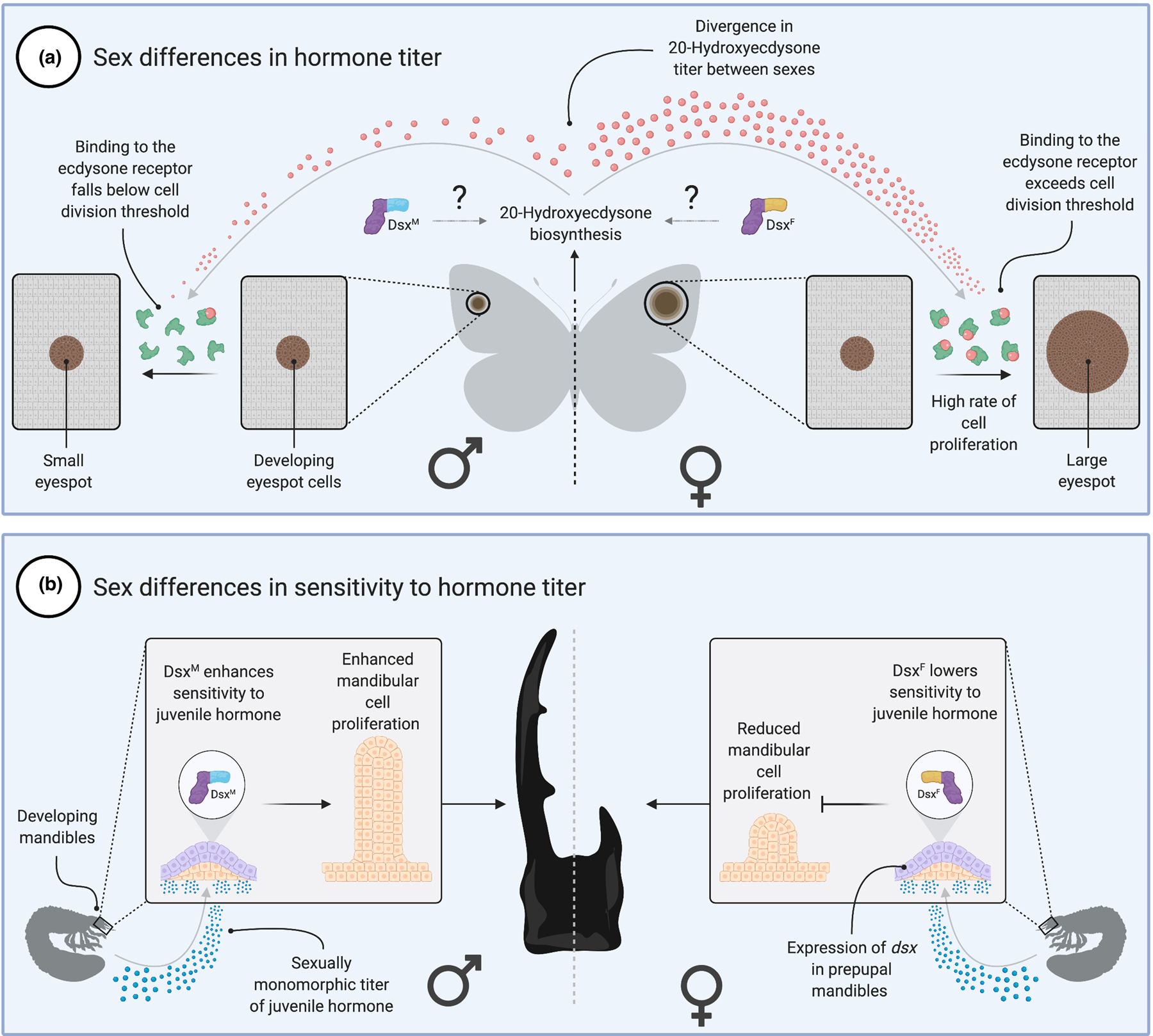
Most animal species consist of two distinct sexes. At the morphological, physiological, and behavioral levels the differences between males and females are numerous and dramatic, yet at the genomic level they are often slight or absent. This disconnect is overcome because simple genetic differences or environmental signals are able to direct the sex-specific expression of a shared genome. A canonical picture of how this process works in insects emerged from decades of work on Drosophila. But recent years have seen an explosion of molecular-genetic and developmental work on a broad range of insects. Drawing these studies together, we describe the evolution of sexual dimorphism from a comparative perspective and argue that insect sex determination and differentiation systems are composites of rapidly evolving and highly conserved elements.
Copyright © 2021 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Conflict of Interest
No conflict of interest exists.
Figures
References
-
- Millington JW, Rideout EJ: Sex differences in Drosophila development and physiology. Curr Opin Physiol 2018, 6:46–56.
-
- Camara N, Whitworth C, Van Doren M: Chapter 3 The Creation of Sexual Dimorphism in the Drosophila Soma. In Current Topics in Developmental Biology. 2008:65–107. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
