

A Translation-Aborting Small Open Reading Frame in the Intergenic Region Promotes Translation of a Mg2+ Transporter in Salmonella Typhimurium
- PMID: 33849981
- PMCID: PMC8092293
- DOI: 10.1128/mBio.03376-20
A Translation-Aborting Small Open Reading Frame in the Intergenic Region Promotes Translation of a Mg2+ Transporter in Salmonella Typhimurium
Abstract
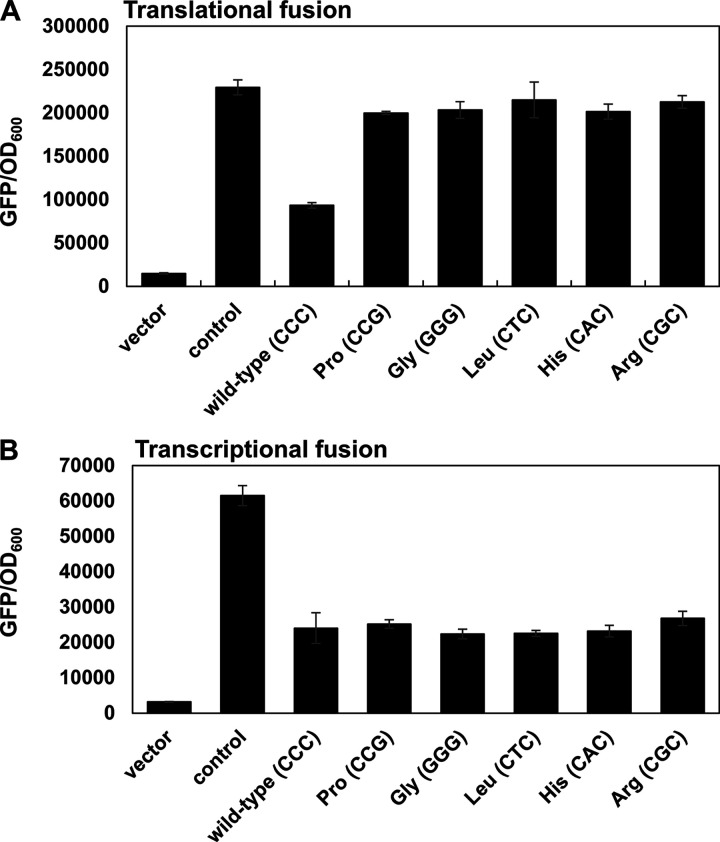
Bacterial mRNAs often harbor upstream open reading frames (uORFs) in the 5' untranslated regions (UTRs). Translation of the uORF usually affects downstream gene expression at the levels of transcription and/or translation initiation. Unlike other uORFs mostly located in the 5' UTR, we discovered an 8-amino-acid ORF, designated mgtQ, in the intergenic region between the mgtC virulence gene and the mgtB Mg2+ transporter gene in the Salmonella mgtCBRU operon. Translation of mgtQ promotes downstream mgtB Mg2+ transporter expression at the level of translation by releasing the ribosome-binding sequence of the mgtB gene that is sequestered in a translation-inhibitory stem-loop structure. Interestingly, mgtQ Asp2 and Glu5 codons that induce ribosome destabilization are required for mgtQ-mediated mgtB translation. Moreover, the mgtQ Asp and Glu codons-mediated mgtB translation is counteracted by the ribosomal subunit L31 that stabilizes ribosome. Substitution of the Asp2 and Glu5 codons in mgtQ decreases MgtB Mg2+ transporter production and thus attenuates Salmonella virulence in mice, likely by limiting Mg2+ acquisition during infection.IMPORTANCE Translation initiation regions in mRNAs that include the ribosome-binding site (RBS) and the start codon are often sequestered within a secondary structure. Therefore, to initiate protein synthesis, the mRNA secondary structure must be unfolded to allow the RBS to be accessible to the ribosome. Such unfolding can be achieved by various mechanisms that include translation of a small upstream open reading frame (uORF). In the intracellular pathogen Salmonella enterica serovar Typhimurium, translation of the Mg2+ transporter mgtB gene is enhanced by an 8-amino-acid upstream ORF, namely, mgtQ, that harbors Asp and Glu codons, which are likely to destabilize ribosome during translation. Translation of the mgtQ ORF promotes the formation of a stem-loop mRNA structure sequestering anti-RBS and thus releases the mgtB RBS. Because mgtQ-mediated MgtB Mg2+ transporter production is required for Salmonella virulence, this pathogen seems to control the virulence determinant production exquisitely via this uORF during infection.
Keywords: intergenic region; ribosome destabilization; translation-inhibitory stem-loop structure; uORF.
Copyright © 2021 Choi et al.
Figures






References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
