

Repeated mutation of a developmental enhancer contributed to human thermoregulatory evolution
- PMID: 33850016
- PMCID: PMC8072367
- DOI: 10.1073/pnas.2021722118
Repeated mutation of a developmental enhancer contributed to human thermoregulatory evolution
Abstract
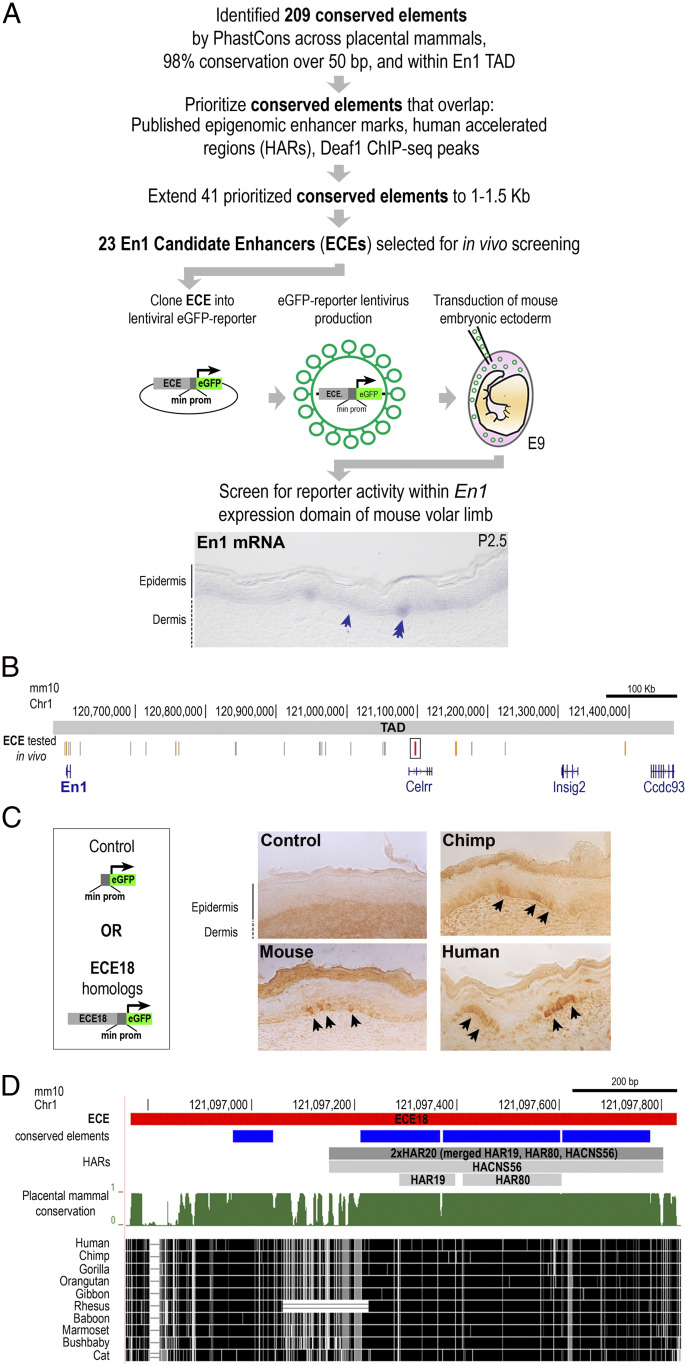
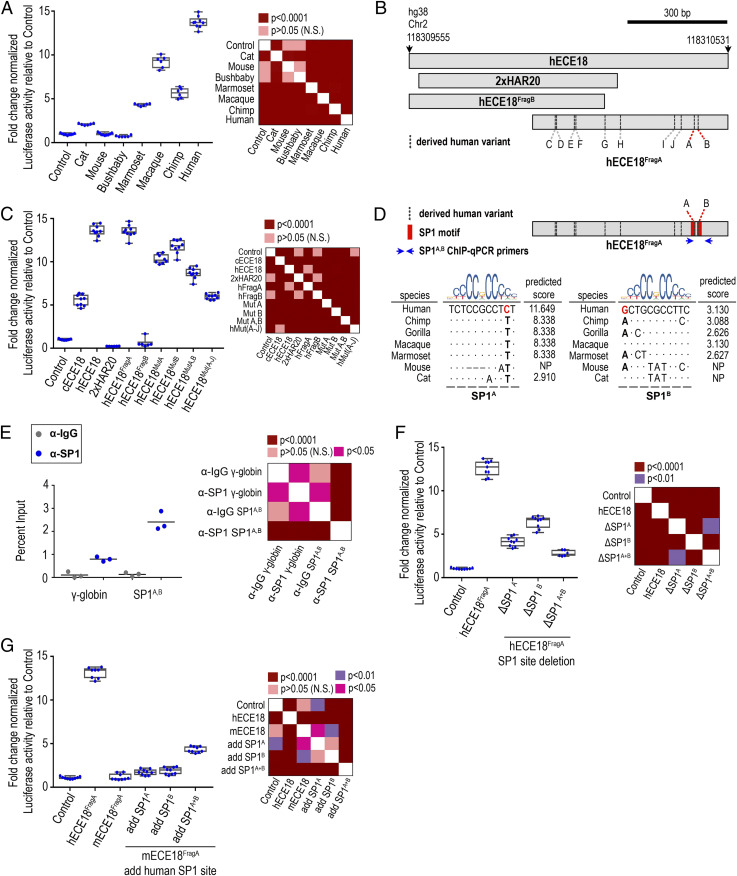
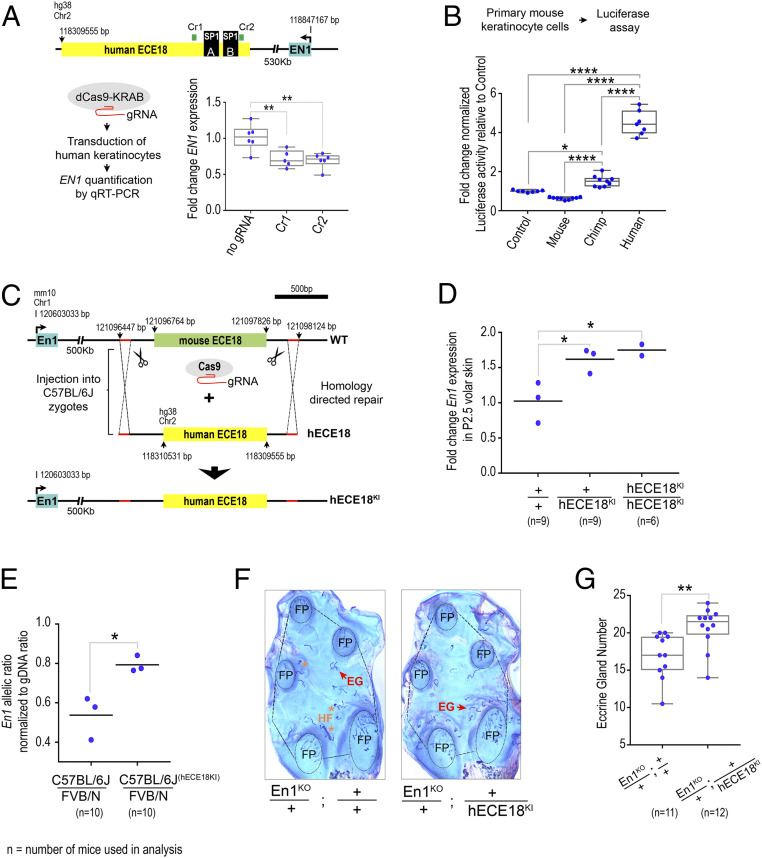
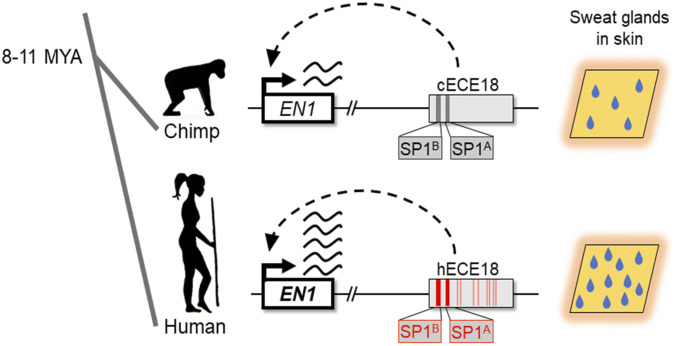
Humans sweat to cool their bodies and have by far the highest eccrine sweat gland density among primates. Humans' high eccrine gland density has long been recognized as a hallmark human evolutionary adaptation, but its genetic basis has been unknown. In humans, expression of the Engrailed 1 (EN1) transcription factor correlates with the onset of eccrine gland formation. In mice, regulation of ectodermal En1 expression is a major determinant of natural variation in eccrine gland density between strains, and increased En1 expression promotes the specification of more eccrine glands. Here, we show that regulation of EN1 has evolved specifically on the human lineage to promote eccrine gland formation. Using comparative genomics and validation of ectodermal enhancer activity in mice, we identified a human EN1 skin enhancer, hECE18. We showed that multiple epistatically interacting derived substitutions in the human ECE18 enhancer increased its activity compared with nonhuman ape orthologs in cultured keratinocytes. Repression of hECE18 in human cultured keratinocytes specifically attenuated EN1 expression, indicating this element positively regulates EN1 in this context. In a humanized enhancer knock-in mouse, hECE18 increased developmental En1 expression in the skin to induce the formation of more eccrine glands. Our study uncovers a genetic basis contributing to the evolution of one of the most singular human adaptations and implicates multiple interacting mutations in a single enhancer as a mechanism for human evolutionary change.
Keywords: Engrailed 1; eccrine gland; human evolution; regulatory evolution; sweat.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Newman R. W., Why man is such a sweaty and thirsty naked animal: A speculative review. Hum. Biol. 42, 12–27 (1970). - PubMed
-
- Kuno Y., Human Perspiration (Thomas, 1956).
-
- Folk G. E. Jr, Semken H. A. Jr, The evolution of sweat glands. Int. J. Biometeorol. 35, 180–186 (1991). - PubMed
-
- Al-Ramamneh D., Gerken M., Riek A., Effect of shearing on water turnover and thermobiological variables in German Blackhead mutton sheep. J. Anim. Sci. 89, 4294–4304 (2011).Corrected in: J. Anim. Sci.90, 395 (2012). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
