

Cardenolides, toxicity, and the costs of sequestration in the coevolutionary interaction between monarchs and milkweeds
- PMID: 33850021
- PMCID: PMC8072370
- DOI: 10.1073/pnas.2024463118
Cardenolides, toxicity, and the costs of sequestration in the coevolutionary interaction between monarchs and milkweeds
Abstract
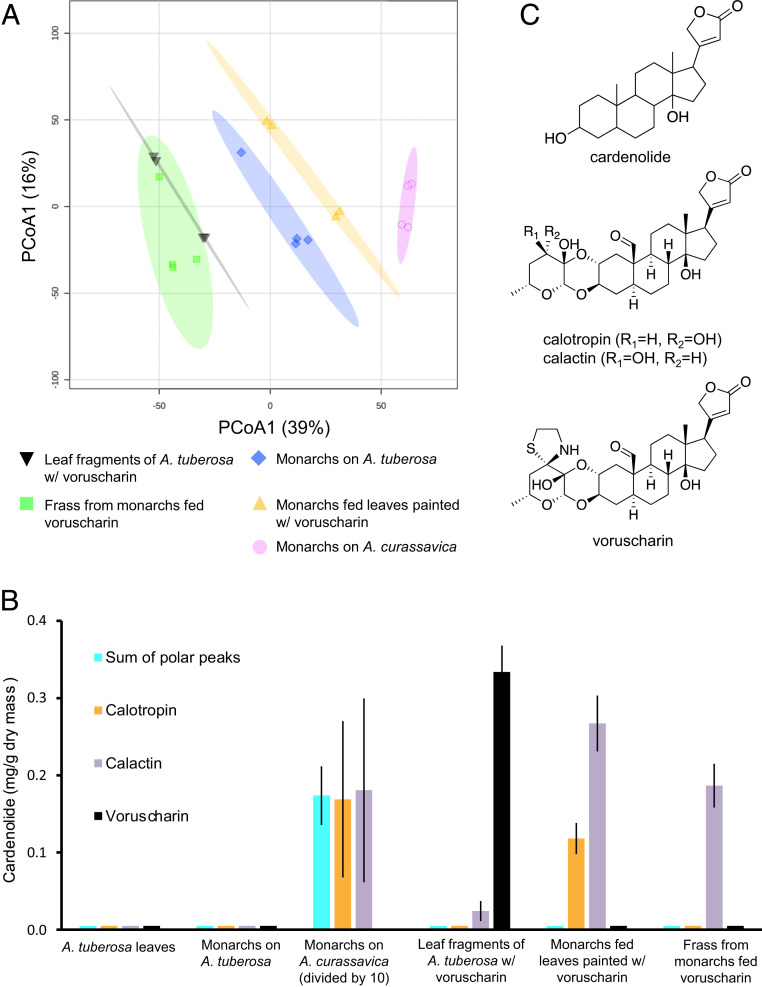
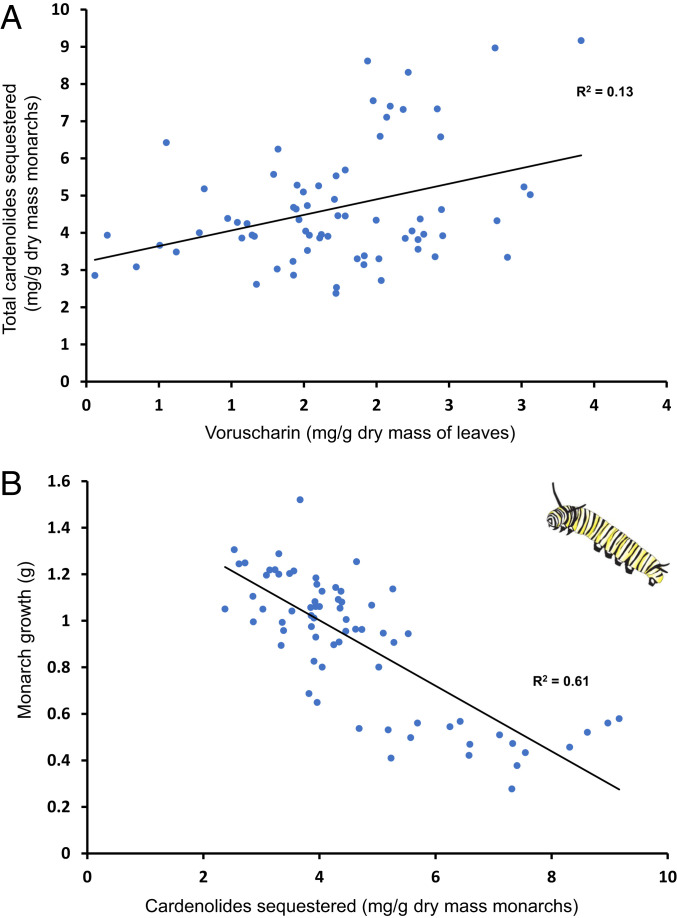
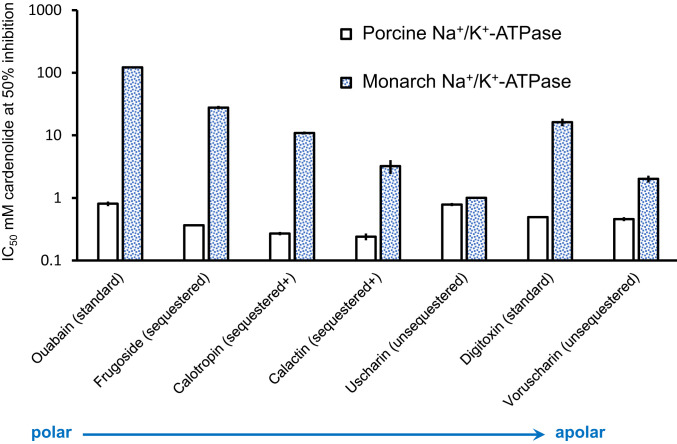
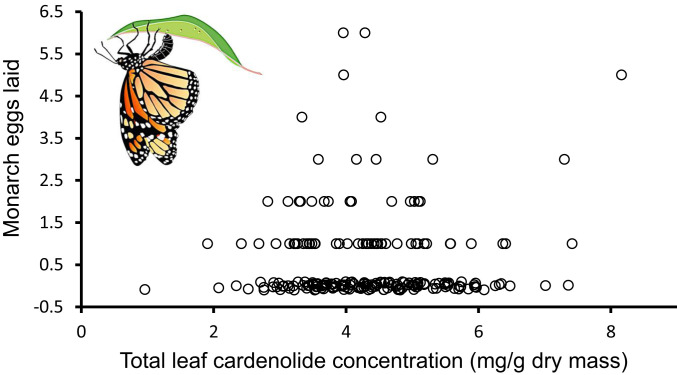
For highly specialized insect herbivores, plant chemical defenses are often co-opted as cues for oviposition and sequestration. In such interactions, can plants evolve novel defenses, pushing herbivores to trade off benefits of specialization with costs of coping with toxins? We tested how variation in milkweed toxins (cardenolides) impacted monarch butterfly (Danaus plexippus) growth, sequestration, and oviposition when consuming tropical milkweed (Asclepias curassavica), one of two critical host plants worldwide. The most abundant leaf toxin, highly apolar and thiazolidine ring-containing voruscharin, accounted for 40% of leaf cardenolides, negatively predicted caterpillar growth, and was not sequestered. Using whole plants and purified voruscharin, we show that monarch caterpillars convert voruscharin to calotropin and calactin in vivo, imposing a burden on growth. As shown by in vitro experiments, this conversion is facilitated by temperature and alkaline pH. We next employed toxin-target site experiments with isolated cardenolides and the monarch's neural Na+/K+-ATPase, revealing that voruscharin is highly inhibitory compared with several standards and sequestered cardenolides. The monarch's typical >50-fold enhanced resistance to cardenolides compared with sensitive animals was absent for voruscharin, suggesting highly specific plant defense. Finally, oviposition was greatest on intermediate cardenolide plants, supporting the notion of a trade-off between benefits and costs of sequestration for this highly specialized herbivore. There is apparently ample opportunity for continued coevolution between monarchs and milkweeds, although the diffuse nature of the interaction, due to migration and interaction with multiple milkweeds, may limit the ability of monarchs to counteradapt.
Keywords: chemical ecology; coevolution; milkweeds Asclepias; monarch butterfly (Danaus plexippus); plant–insect interactions.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Cornell H. V., Hawkins B. A., Herbivore responses to plant secondary compounds: A test of phytochemical coevolution theory. Am. Nat. 161, 507–522 (2003). - PubMed
-
- Rothwell E. M., Holeski L. M., Phytochemical defences and performance of specialist and generalist herbivores: A meta‐analysis. Ecol. Entomol. 45, 396–405 (2019).
-
- Berenbaum M. R., Postgenomic chemical ecology: From genetic code to ecological interactions. J. Chem. Ecol. 28, 873–896 (2002). - PubMed
-
- Renwick J. A. A., Haribal M., Gouinguené S., Städler E., Isothiocyanates stimulating oviposition by the diamondback moth, Plutella xylostella. J. Chem. Ecol. 32, 755–766 (2006). - PubMed
-
- Nieminen M., Suomi J., Van Nouhuys S., Sauri P., Riekkola M.-L., Effect of iridoid glycoside content on oviposition host plant choice and parasitism in a specialist herbivore. J. Chem. Ecol. 29, 823–844 (2003). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
