

Avid binding by B cells to the Plasmodium circumsporozoite protein repeat suppresses responses to protective subdominant epitopes
- PMID: 33852850
- PMCID: PMC8052187
- DOI: 10.1016/j.celrep.2021.108996
Avid binding by B cells to the Plasmodium circumsporozoite protein repeat suppresses responses to protective subdominant epitopes
Abstract
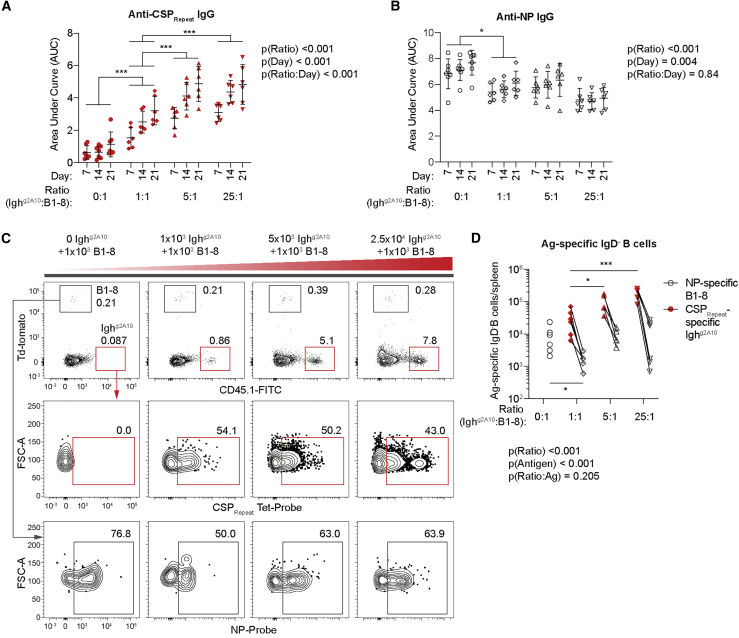
Antibodies targeting the NANP/NVDP repeat domain of the Plasmodium falciparum circumsporozoite protein (CSPRepeat) can protect against malaria. However, it has also been suggested that the CSPRepeat is a decoy that prevents the immune system from mounting responses against other domains of CSP. Here, we show that, following parasite immunization, B cell responses to the CSPRepeat are immunodominant over responses to other CSP domains despite the presence of similar numbers of naive B cells able to bind these regions. We find that this immunodominance is driven by avid binding of the CSPRepeat to cognate B cells that are able to expand at the expense of B cells with other specificities. We further show that mice immunized with repeat-truncated CSP molecules develop responses to subdominant epitopes and are protected against malaria. These data demonstrate that the CSPRepeat functions as a decoy, but truncated CSP molecules may be an approach for malaria vaccination.
Keywords: B cells; Plasmodium falciparum; circumsporozoite protein; immunodominance; immunodomination; malaria; malaria vaccines.
Copyright © 2021 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures







References
-
- Abbott R.K., Lee J.H., Menis S., Skog P., Rossi M., Ota T., Kulp D.W., Bhullar D., Kalyuzhniy O., Havenar-Daughton C. Precursor Frequency and Affinity Determine B Cell Competitive Fitness in Germinal Centers, Tested with Germline-Targeting HIV Vaccine Immunogens. Immunity. 2018;48:133–146.e6. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
