

Convergence of a common solution for broad ebolavirus neutralization by glycan cap-directed human antibodies
- PMID: 33852862
- PMCID: PMC8133395
- DOI: 10.1016/j.celrep.2021.108984
Convergence of a common solution for broad ebolavirus neutralization by glycan cap-directed human antibodies
Abstract
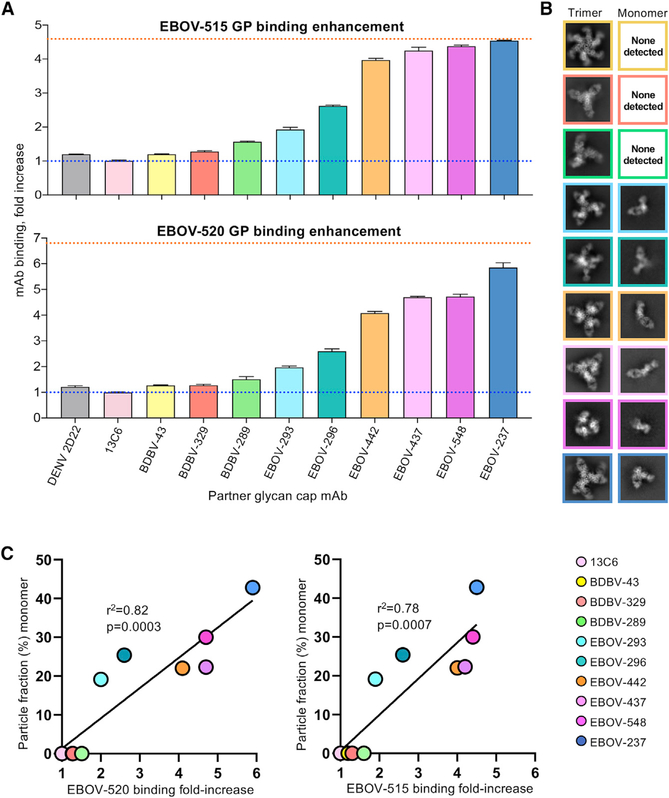
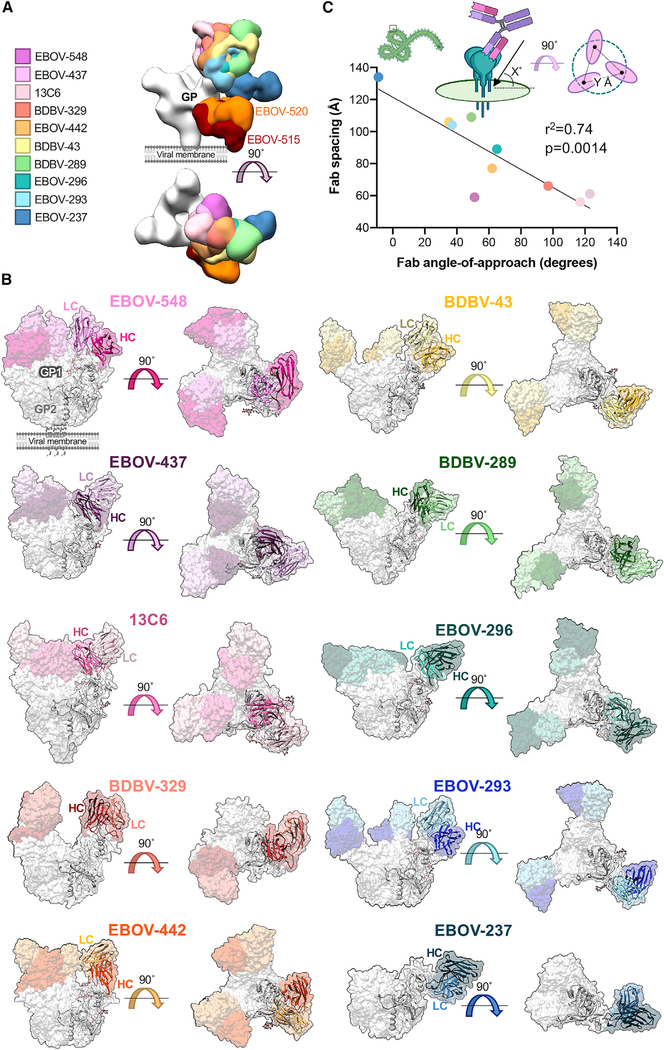
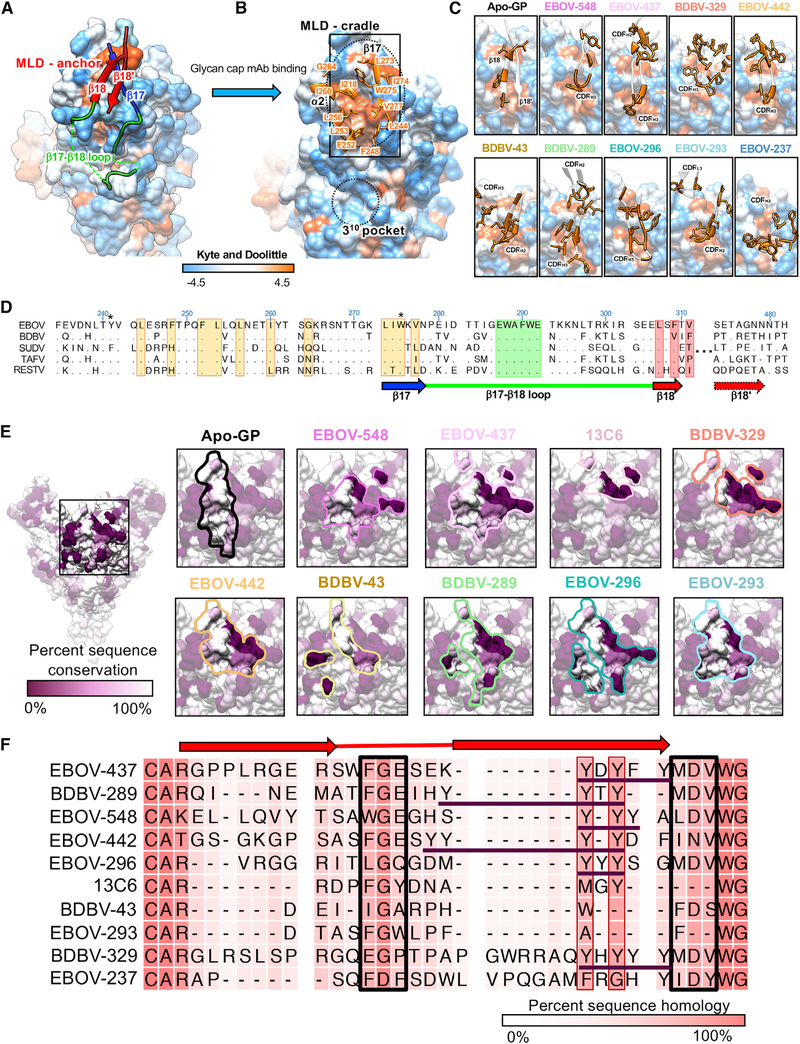
Antibodies that target the glycan cap epitope on the ebolavirus glycoprotein (GP) are common in the adaptive response of survivors. A subset is known to be broadly neutralizing, but the details of their epitopes and basis for neutralization are not well understood. Here, we present cryoelectron microscopy (cryo-EM) structures of diverse glycan cap antibodies that variably synergize with GP base-binding antibodies. These structures describe a conserved site of vulnerability that anchors the mucin-like domains (MLDs) to the glycan cap, which we call the MLD anchor and cradle. Antibodies that bind to the MLD cradle share common features, including use of IGHV1-69 and IGHJ6 germline genes, which exploit hydrophobic residues and form β-hairpin structures to mimic the MLD anchor, disrupt MLD attachment, destabilize GP quaternary structure, and block cleavage events required for receptor binding. Our results provide a molecular basis for ebolavirus neutralization by broadly reactive glycan cap antibodies.
Keywords: Ebola virus; antibody; antibody therapeutics; broadly neutralizing; ebolaviruses; filoviruses; glycan cap; mAb.
Copyright © 2021 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests A.L.B., E.D., and B.J.D. are employees of Integral Molecular. B.J.D. is a shareholder of Integral Molecular. J.E.C. has served as a consultant for Lilly and Luna Biologics, is a member of the Scientific Advisory Boards of CompuVax and Meissa Vaccines, and is the founder of IDBiologics. The Crowe laboratory at Vanderbilt University Medical Center has received sponsored research agreements from and IDBiologics and AstraZeneca. Vanderbilt University has applied for a patent that is related to antibodies discussed in this work. All other authors declare no competing interests.
Figures

Similar articles
-
Cross-reactive neutralizing human survivor monoclonal antibody BDBV223 targets the ebolavirus stalk.Nat Commun. 2019 Apr 17;10(1):1788. doi: 10.1038/s41467-019-09732-7. Nat Commun. 2019. PMID: 30996276 Free PMC article.
-
Structural and molecular basis for Ebola virus neutralization by protective human antibodies.Science. 2016 Mar 18;351(6279):1343-6. doi: 10.1126/science.aad6117. Epub 2016 Feb 25. Science. 2016. PMID: 26917592 Free PMC article.
-
Multifunctional Pan-ebolavirus Antibody Recognizes a Site of Broad Vulnerability on the Ebolavirus Glycoprotein.Immunity. 2018 Aug 21;49(2):363-374.e10. doi: 10.1016/j.immuni.2018.06.018. Epub 2018 Jul 17. Immunity. 2018. PMID: 30029854 Free PMC article.
-
Structural Biology Illuminates Molecular Determinants of Broad Ebolavirus Neutralization by Human Antibodies for Pan-Ebolavirus Therapeutic Development.Front Immunol. 2022 Jan 10;12:808047. doi: 10.3389/fimmu.2021.808047. eCollection 2021. Front Immunol. 2022. PMID: 35082794 Free PMC article. Review.
-
Structural basis for differential neutralization of ebolaviruses.Viruses. 2012 Apr;4(4):447-70. doi: 10.3390/v4040447. Epub 2012 Apr 5. Viruses. 2012. PMID: 22590681 Free PMC article. Review.
Cited by
-
Discovery of Nanosota-EB1 and -EB2 as Novel Nanobody Inhibitors Against Ebola Virus Infection.PLoS Pathog. 2024 Dec 23;20(12):e1012817. doi: 10.1371/journal.ppat.1012817. eCollection 2024 Dec. PLoS Pathog. 2024. PMID: 39715280 Free PMC article.
-
Protocol for quantification of intercellular connection transmission in ebolavirus infections using ImageJ.STAR Protoc. 2024 Dec 20;5(4):103363. doi: 10.1016/j.xpro.2024.103363. Epub 2024 Oct 5. STAR Protoc. 2024. PMID: 39369387 Free PMC article.
-
Glycan shield of the ebolavirus envelope glycoprotein GP.Commun Biol. 2022 Aug 4;5(1):785. doi: 10.1038/s42003-022-03767-1. Commun Biol. 2022. PMID: 35927436 Free PMC article.
-
Computational mining of B cell receptor repertoires reveals antigen-specific and convergent responses to Ebola vaccination.Front Immunol. 2024 Jul 8;15:1383753. doi: 10.3389/fimmu.2024.1383753. eCollection 2024. Front Immunol. 2024. PMID: 39040106 Free PMC article. Clinical Trial.
-
Pan-ebolavirus protective therapy by two multifunctional human antibodies.Cell. 2021 Oct 28;184(22):5593-5607.e18. doi: 10.1016/j.cell.2021.09.035. Cell. 2021. PMID: 34715022 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
