

The connectome of neural crest enhancers reveals regulatory features of signaling systems
- PMID: 33852891
- PMCID: PMC8445387
- DOI: 10.1016/j.devcel.2021.03.024
The connectome of neural crest enhancers reveals regulatory features of signaling systems
Abstract
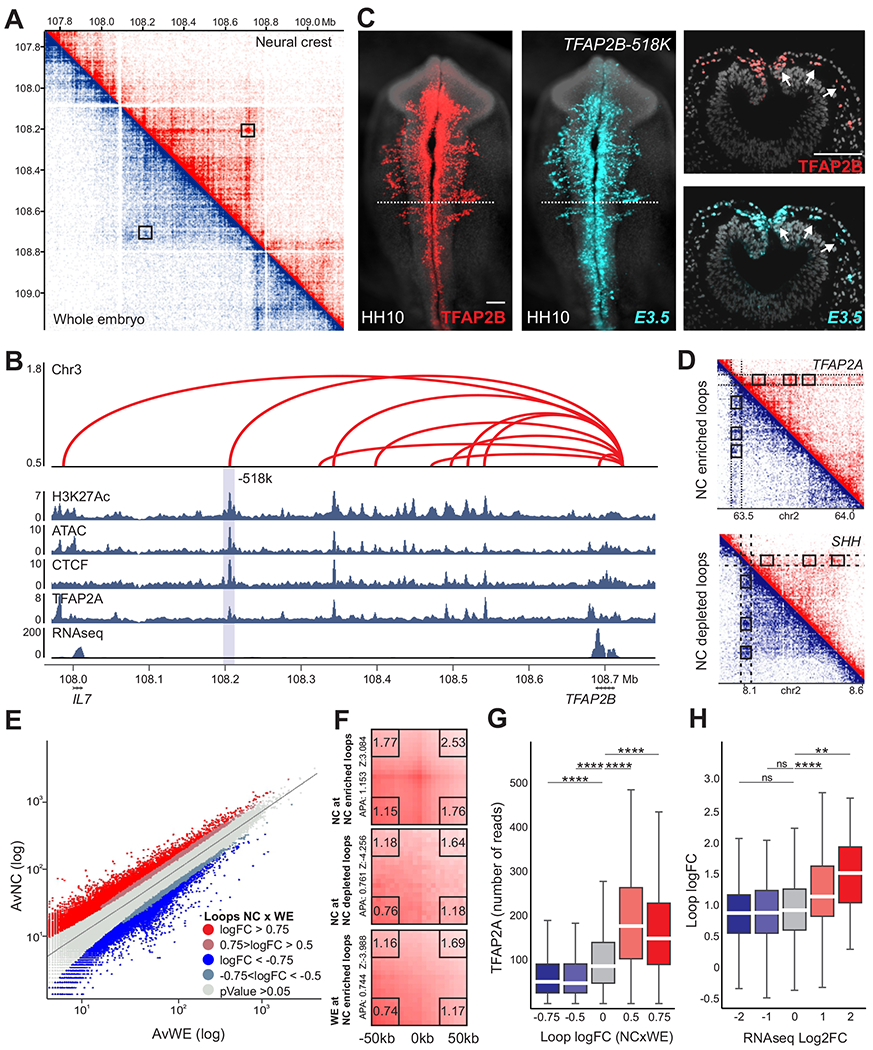
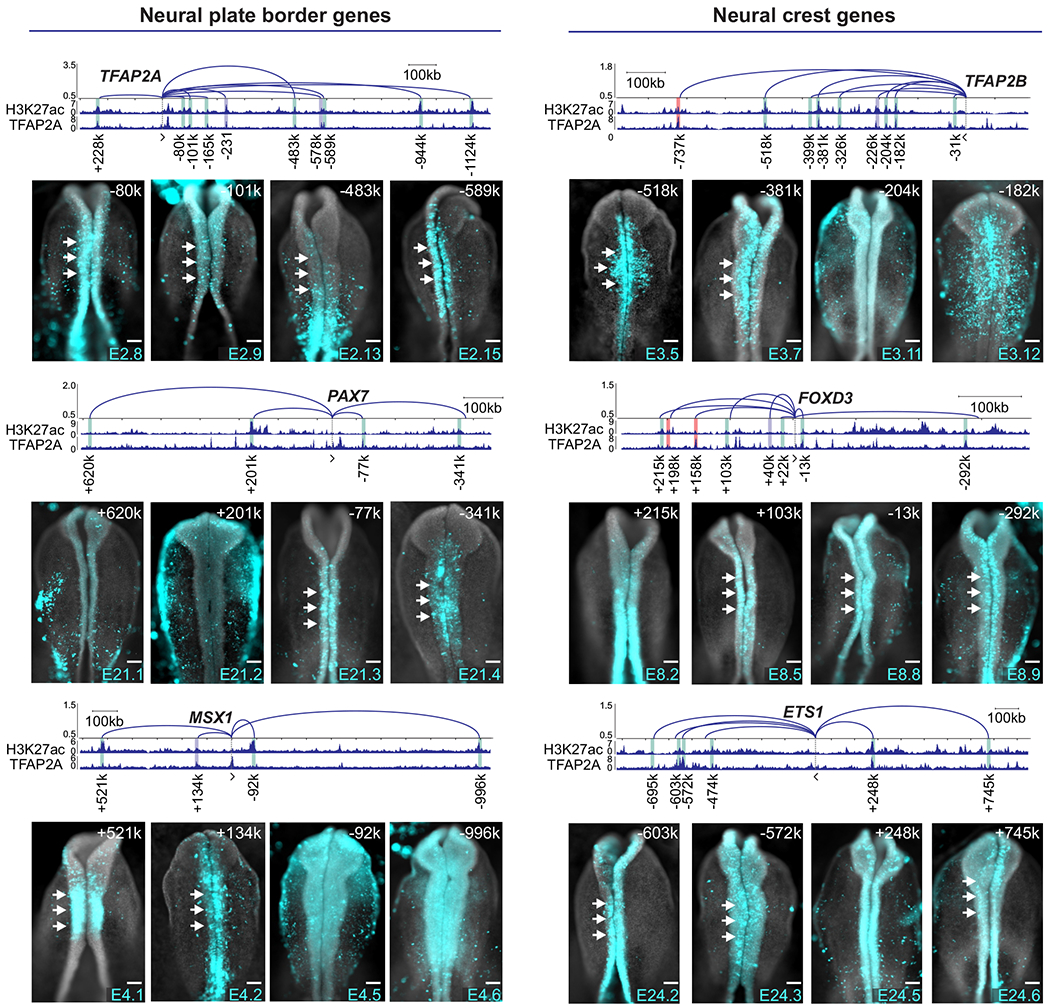
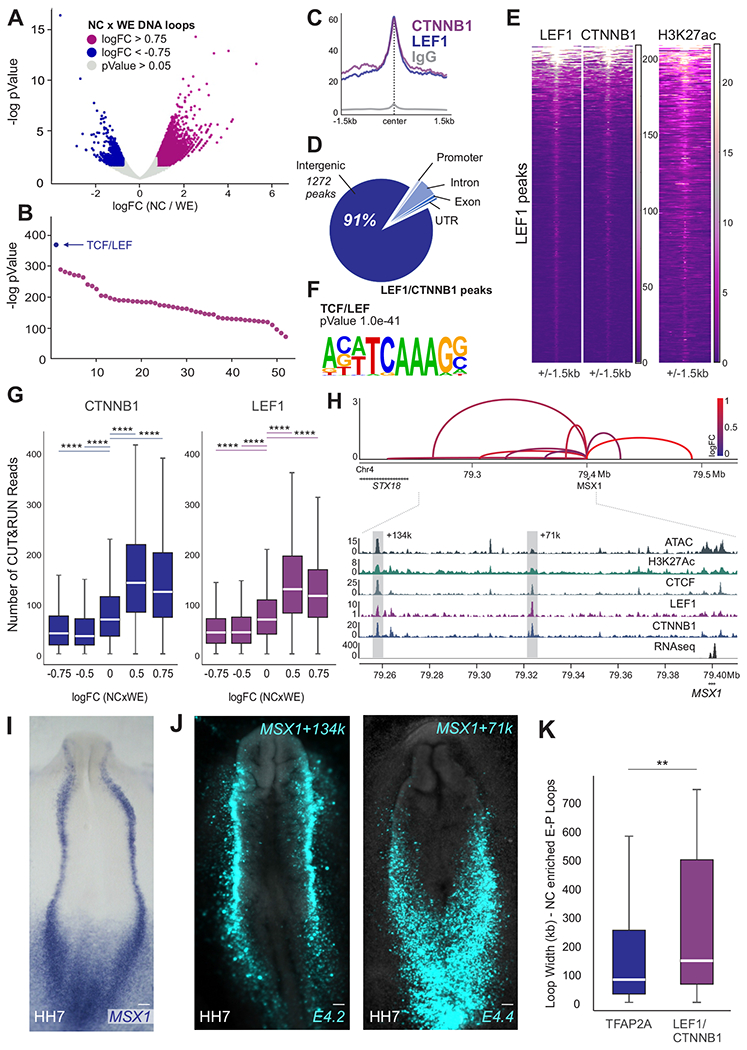
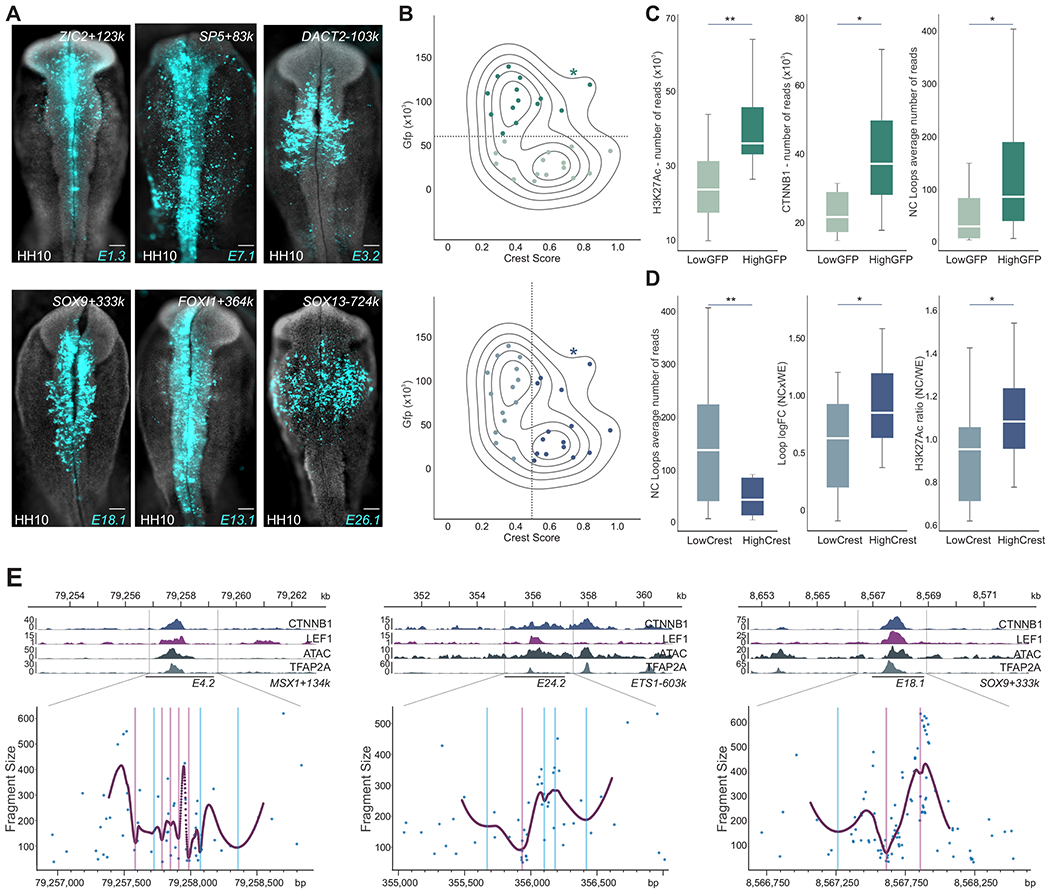
Cell fate commitment is controlled by cis-regulatory elements often located in remote regions of the genome. To examine the role of long-range DNA interactions in early development, we generated a high-resolution contact map of active enhancers in avian neural crest cells. This analysis uncovered a diverse repertoire of enhancers that are part of the gene regulatory network underlying specification. We found that neural crest identity is largely regulated by cis-regulatory elements that propagate signaling inputs to network components. These genomic sensors display a combination of optimal and suboptimal TCF/LEF-binding sites, which allow cells to respond to Wnt signaling in a position-dependent manner. We propose that, rather than acting as upstream activators, signaling systems feed into regulatory circuits in a hub-and-spoke architecture. These results shed light on the tridimensional organization of the neural crest genome and define how signaling systems provide progenitors with spatial cues that transform their molecular identity.
Keywords: Hi-ChIP; Wnt; Wnt signaling; enhancer connectome; enhancers; neural crest cells; nuclear architecture; signaling systems.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- AMANO T, SAGAI T, TANABE H, MIZUSHINA Y, NAKAZAWA H & SHIROISHI T 2009. Chromosomal dynamics at the Shh locus: limb bud-specific differential regulation of competence and active transcription. Dev Cell, 16, 47–57. - PubMed
-
- BARON R & KNEISSEL M 2013. WNT signaling in bone homeostasis and disease: from human mutations to treatments. Nat Med, 19, 179–92. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
