

Loss of LUC7L2 and U1 snRNP subunits shifts energy metabolism from glycolysis to OXPHOS
- PMID: 33852893
- PMCID: PMC8314041
- DOI: 10.1016/j.molcel.2021.02.033
Loss of LUC7L2 and U1 snRNP subunits shifts energy metabolism from glycolysis to OXPHOS
Abstract
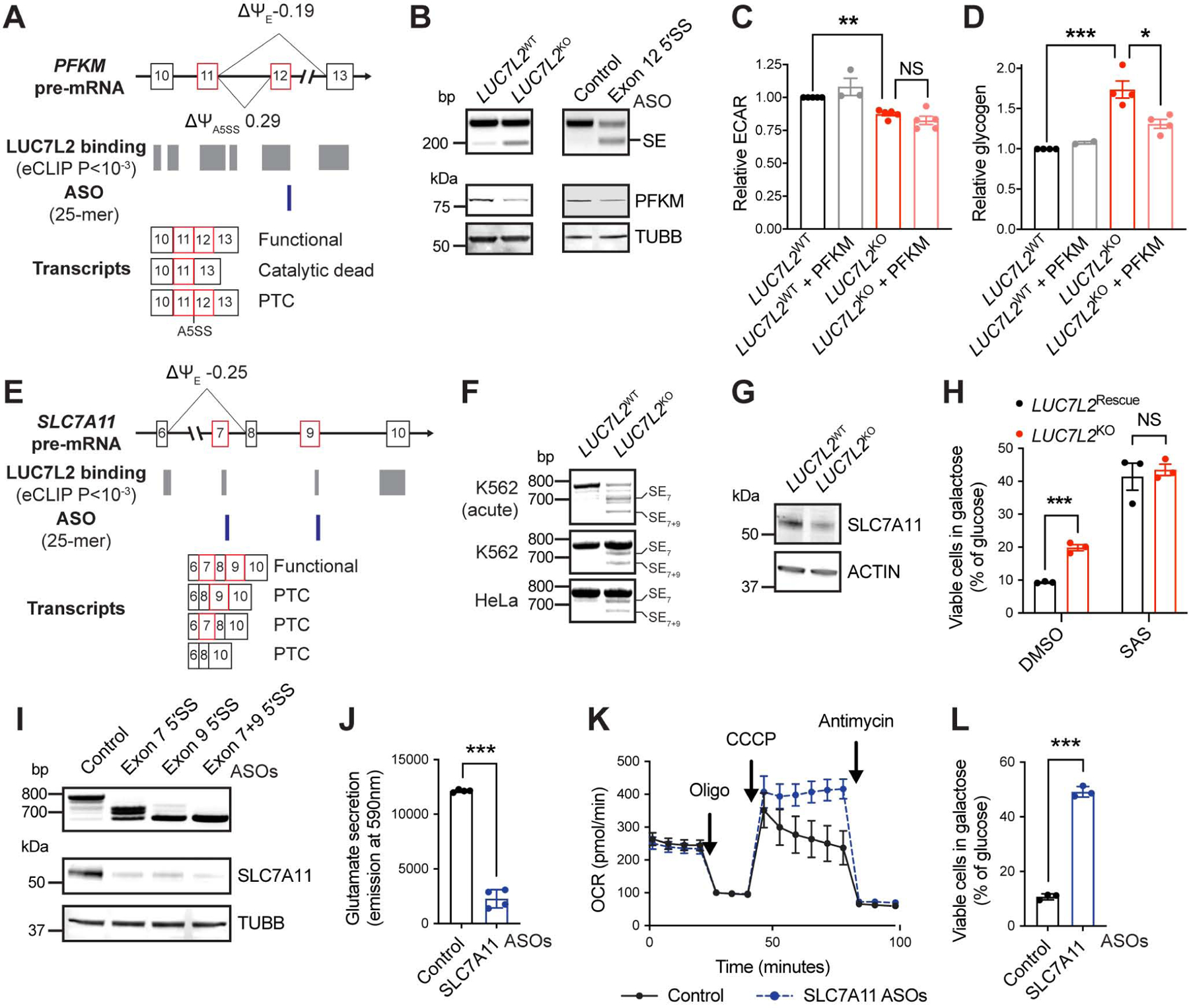
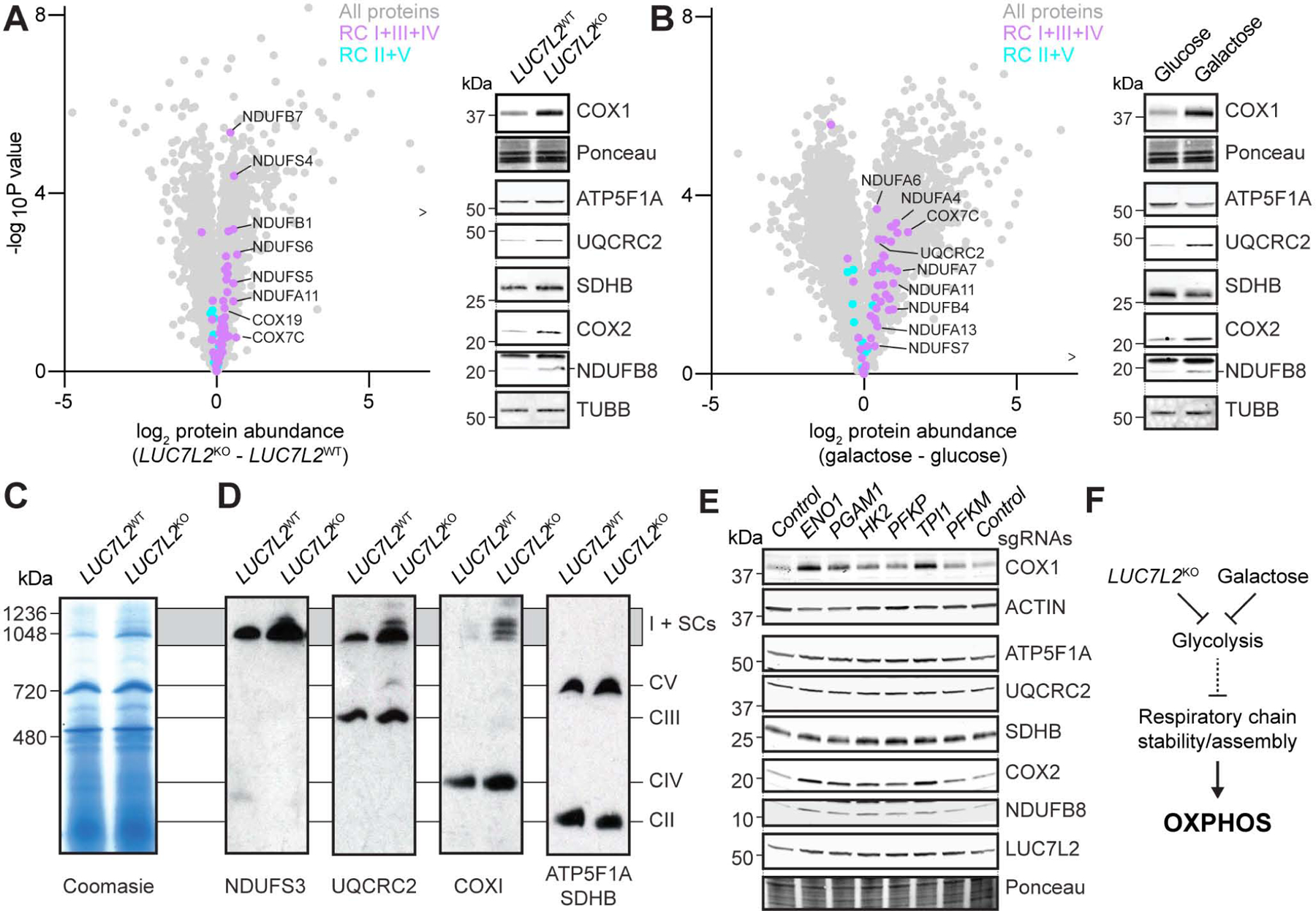
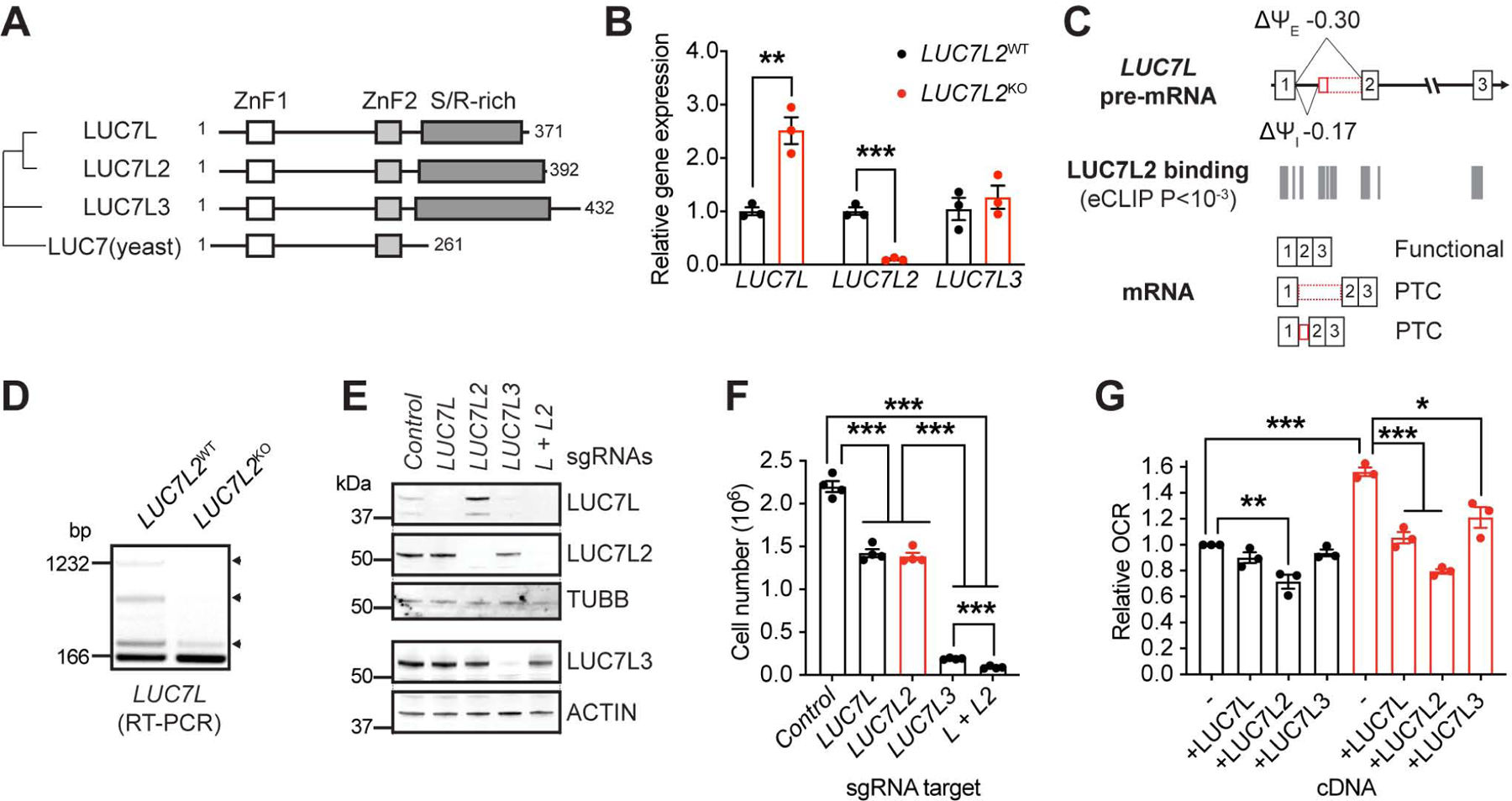
Oxidative phosphorylation (OXPHOS) and glycolysis are the two major pathways for ATP production. The reliance on each varies across tissues and cell states, and can influence susceptibility to disease. At present, the full set of molecular mechanisms governing the relative expression and balance of these two pathways is unknown. Here, we focus on genes whose loss leads to an increase in OXPHOS activity. Unexpectedly, this class of genes is enriched for components of the pre-mRNA splicing machinery, in particular for subunits of the U1 snRNP. Among them, we show that LUC7L2 represses OXPHOS and promotes glycolysis by multiple mechanisms, including (1) splicing of the glycolytic enzyme PFKM to suppress glycogen synthesis, (2) splicing of the cystine/glutamate antiporter SLC7A11 (xCT) to suppress glutamate oxidation, and (3) secondary repression of mitochondrial respiratory supercomplex formation. Our results connect LUC7L2 expression and, more generally, the U1 snRNP to cellular energy metabolism.
Keywords: 7q-; LUC7; MDS; Tarui disease; cancer; ferroptosis; myelodysplastic syndrome; phosphofructokinase; spliceosome; system X(c)(−).
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests V.K.M. is a paid scientific advisor to 5AM Ventures and Janssen Pharmaceuticals. O.S.S. is a paid consultant for Proteinaceous. R.S. holds equity in BlueBird Bio. G.W.Y. is co-founder, member of the Board of Directors, on the scientific advisory board, equity holder, and paid consultant for Locanabio and Eclipse Bioinnovations. G.W.Y. is a visiting professor at the National University of Singapore. G.W.Y.’s interest(s) have been reviewed and approved by the University of California, San Diego in accordance with its conflict of interest policies. A.A.J. and V.K.M. are co-inventors on a US provisional patent application related to the work in this manuscript. The authors declare no other competing interests.
Figures
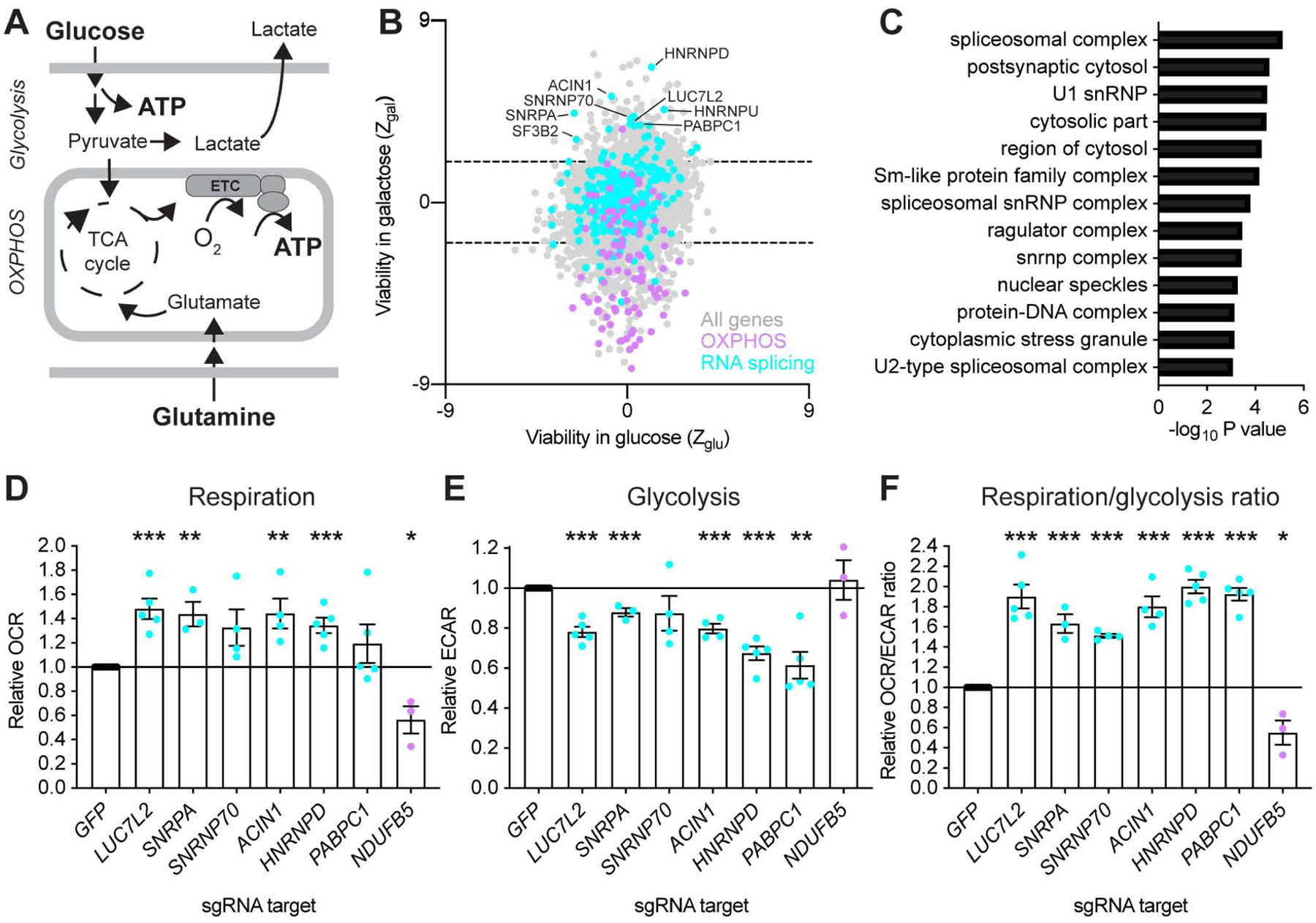
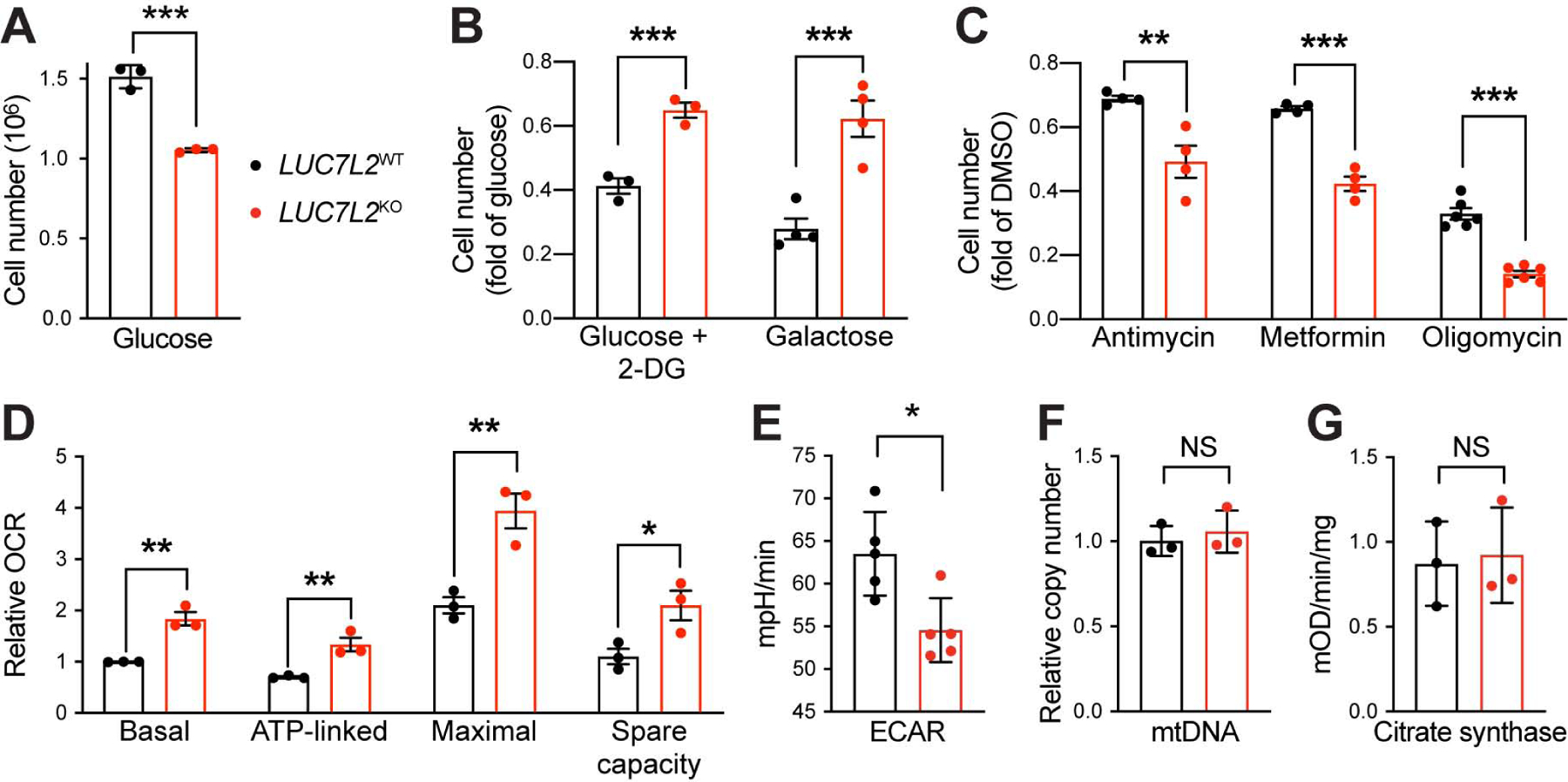
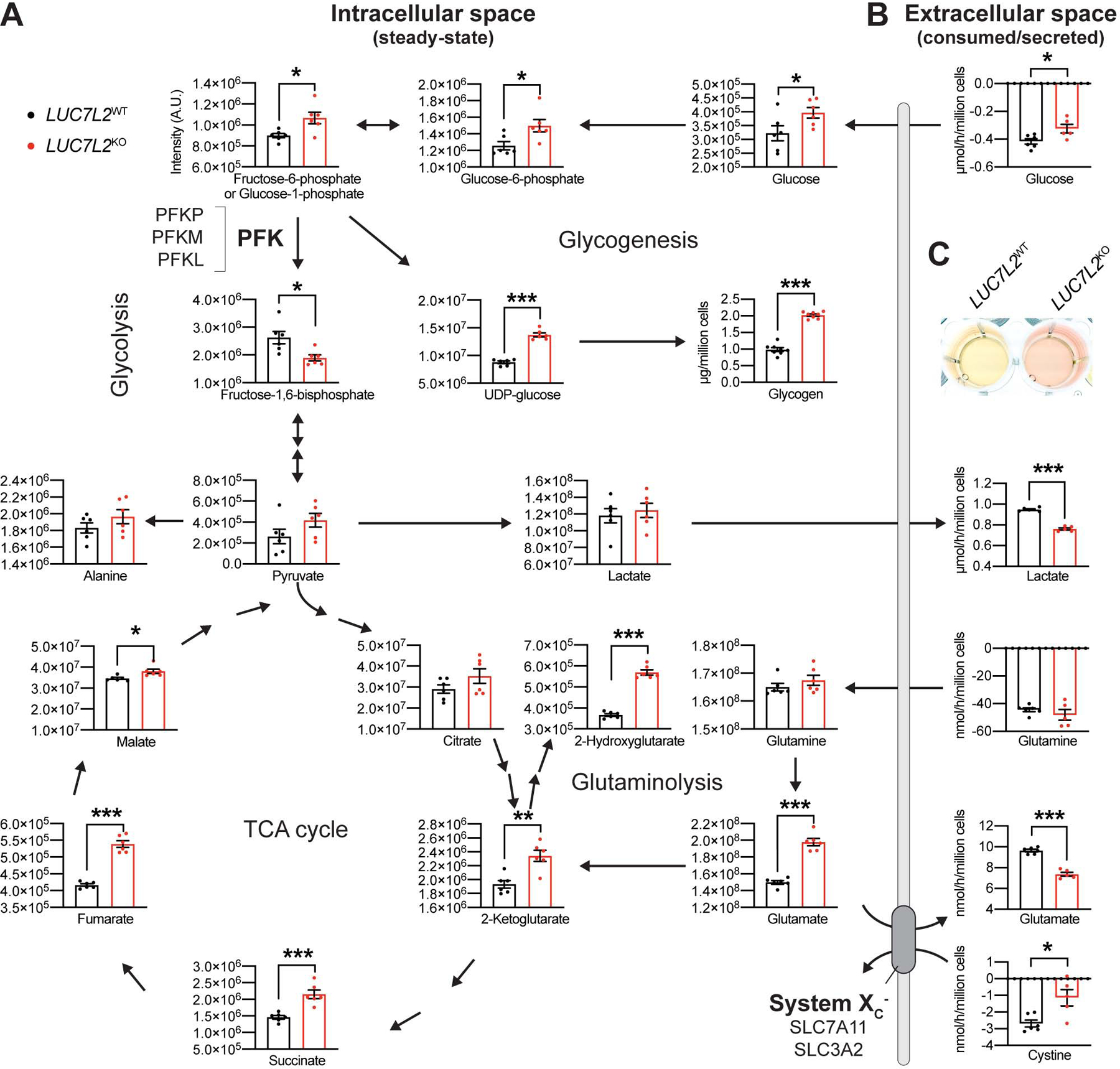
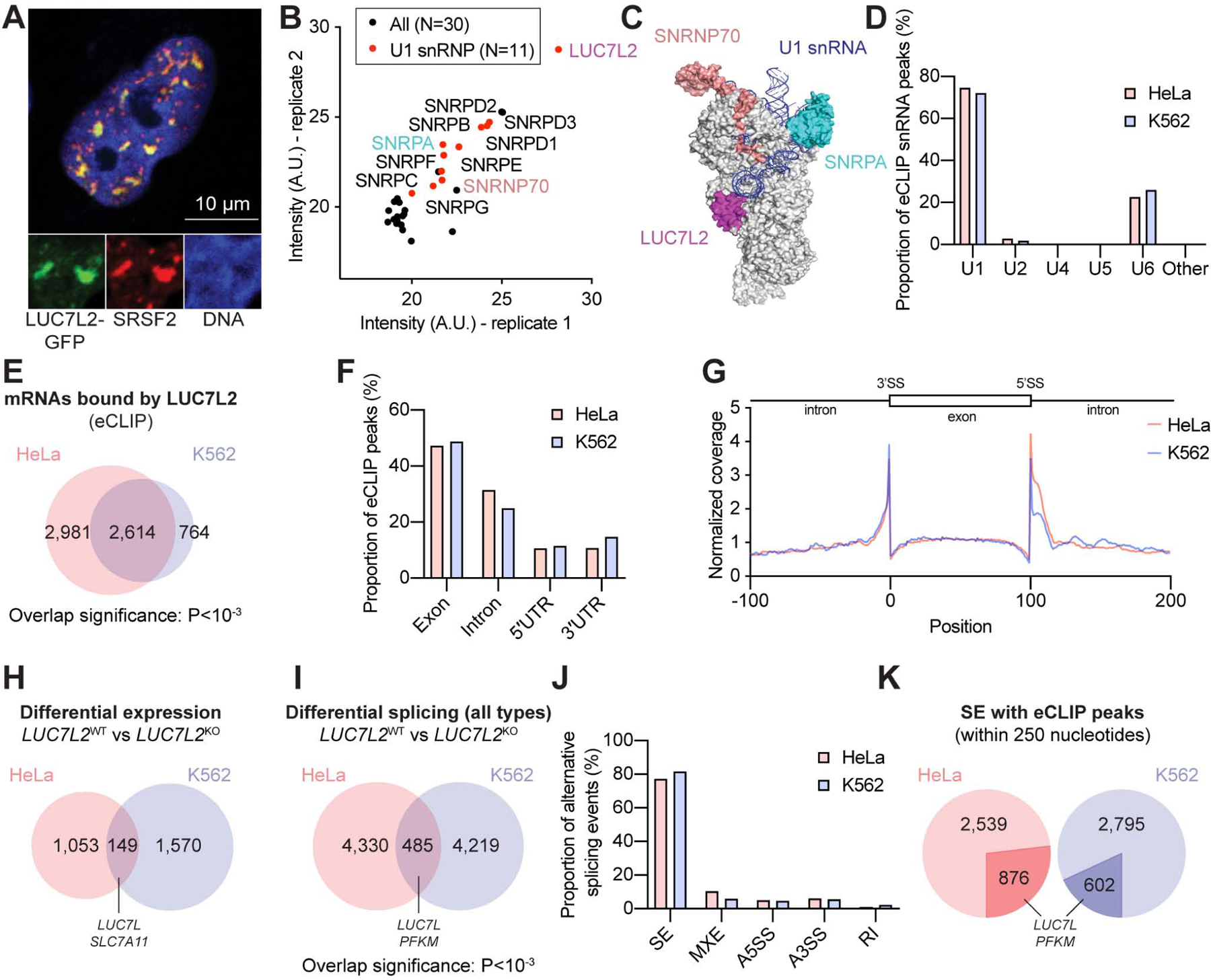
Comment in
-
When cells are down on their LUC7L2, alternative splicing rewires metabolism for OXPHOS.Mol Cell. 2021 May 6;81(9):1859-1860. doi: 10.1016/j.molcel.2021.04.003. Mol Cell. 2021. PMID: 33961773
References
-
- BADGLEY MA, KREMER DM, MAURER HC, DELGIORNO KE, LEE HJ, PUROHIT V, SAGALOVSKIY IR, MA A, KAPILIAN J, FIRL CEM, DECKER AR, SASTRA SA, PALERMO CF, ANDRADE LR, SAJJAKULNUKIT P, ZHANG L, TOLSTYKA ZP, HIRSCHHORN T, LAMB C, LIU T, GU W, SEELEY ES, STONE E, GEORGIOU G, MANOR U, IUGA A, WAHL GM, STOCKWELL BR, LYSSIOTIS CA & OLIVE KP 2020. Cysteine depletion induces pancreatic tumor ferroptosis in mice. Science, 368, 85–89. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
